

Summary
Coronavirus disease 2019 (Covid‐19) is caused by severe acute respiratory syndrome‐coronavirus‐2 (SARS‐CoV‐2) which is responsible for a global pandemic that started in late 2019 in Wuhan, China. To prevent the worldwide spread of this highly pathogenic virus, development of an effective and safe vaccine is urgently needed. The SARS‐CoV‐2 and SARS‐CoV share a high degree of genetic and pathologic identity and share safety and immune‐enhancement concerns regarding vaccine development. Prior animal studies with first generation (whole virus‐based) preparations of SARS‐CoV vaccines (inactivated and attenuated vaccine modalities) indicated the possibility of increased infectivity or eosinophilic infiltration by immunization. Therefore, development of second and third generation safer vaccines (by using modern vaccine platforms) is actively sought for this viral infection. The spike (S) protein of SARS‐CoVs is the main determinant of cell entry and tropism and is responsible for facilitating zoonosis into humans and sustained person‐to‐person transmission. Furthermore, ‘S’ protein contains multiple neutralizing epitopes that play an essential role in the induction of neutralizing antibodies (nAbs) and protective immunity. Moreover, T‐cell responses against the SARS‐CoV‐2 ‘S’ protein have also been characterized that correlate to the IgG and IgA antibody titres in Covid‐19 patients. Thus, S protein is an obvious candidate antigen for inclusion into vaccine platforms against SARS‐CoV‐2 viral infection. This manuscript reviews different characteristics of S protein, its potency and ‘state of the art’ of the vaccine development strategies and platforms using this antigen, for construction of a safe and effective SARS‐CoV‐2 vaccine.
Keywords: RBD, SARS‐CoV‐2, spike, vaccine
Abbreviations
- AAV
adeno‐associated virus
- ACE2
angiotensin‐converting enzyme 2
- ADE
antibody‐dependent enhancement
- ALI
acute lung injury
- ARDS
acute respiratory distress syndrome
- CoV
coronaviruses
- Covid‐19
coronavirus disease 2019
- Cryo‐EM
cryo‐electron microscopy
- E
envelope
- FP
fusion peptide
- GMT
geometric mean titres
- HR
heptad repeat
- HSR
hypersensitivity reaction
- IM
intramuscular
- M
membrane
- mAb
monoclonal antibody
- MERS‐CoV
Middle East respiratory syndrome‐coronavirus
- MNA
microneedle arrays
- MVA
modified vaccinia virus Ankara
- N
nucleocapsid
- nAb
neutralizing antibody
- NK
natural killers
- NSP
non‐structural protein
- NTD
N‐terminal domain
- PBMCs
peripheral blood mononuclear cells
- RBD
receptor‐binding domain
- RCT
randomized clinical trials
- S
spike
- SARS‐CoV‐2
severe acute respiratory syndrome‐coronavirus‐2
- TLR
Toll‐like receptor
- tPA
tissue plasminogen activator
- VH
variable heavy
- VP
virus particle
1. INTRODUCTION
Coronaviruses (CoVs) are enveloped viruses of the Coronaviridae family, composed of a large single‐strand, positive‐sense RNA genome (ranging from 26 to 32 kilobases in length) with 5′‐cap and 3′ poly‐A tail. 1 While some members of the Coronaviridae family might cause mild respiratory symptoms (229E, OC43, NL63 and HKU1), three members including severe acute respiratory syndrome‐CoV (SARS‐CoV), Middle East respiratory syndrome‐CoV (MERS‐CoV) and the novel coronavirus (SARS‐CoV‐2) are highly pathogenic in humans. 2 , 3 SARS‐CoV emerged in China in 2002 and MERS‐CoV in Saudi Arabia in 2012. 4 , 5 , 6 , 7 , 8 The novel coronavirus (2019‐nCoV), first detected in patients with pneumonia in Wuhan, China, in December 2019, 3 , 9 , 10 was named SARS‐CoV‐2 and the disease caused by SARS‐CoV‐2 was named Coronavirus Disease 2019 (Covid‐19). 3 , 11 SARS‐CoV‐2 is believed to have originated from bats, but pangolins are proposed as possible intermediate hosts. 12 Although the mortality rates in SARS‐CoV‐2 infection are not as high compared to that of the SARS‐CoV and MERS‐CoV, SARS‐CoV‐2 is more transmissible and so has claimed considerably more lives. 1 Additionally, the newly reported D614G amino acid change in spike (S) protein seems to have augmented infectivity of the SARS‐CoV‐2. 13 Development of a vaccine against this viral infection is the priority of WHO and other global healthcare organizations. However, several drugs are being evaluated for efficacy in treating SARS‐CoV‐2, among which remdesivir and dexamethasone have shown improved outcomes in very ill patients. 14 , 15 The results of recent randomized clinical trials (RCTs) demonstrated that remdesivir (that received FDA authorization of emergency use in severe Covid‐19 patients) 16 , 17 , 18 and dexamethasone 19 can decrease the recovery time for Covid‐19 hospitalized‐patients under supplemental, oxygen therapy. There are also ongoing RCTs evaluating the safety and efficacy of the immuno‐modulator interferon beta‐1a alone (NCT04385095) or in combination with remdesivir (NCT04492475).
The SARS‐CoV‐2 genome encodes several non‐structural (NSP1‐NSP10 and NSP12‐16) and accessory proteins as well as four structural proteins, including spike (S), envelope (E), membrane (M) and nucleocapsid (N; Figure 1a). Among structural proteins, S, is responsible for binding to cellular‐angiotensin‐converting enzyme 2 (ACE2; which acts as the cellular receptor) and thus is an obvious candidate antigen for vaccine development based on induction of neutralizing antibodies (nAbs) against the virus. 11 , 20
FIGURE 1.

Genome composition of SARS‐CoV‐2. (a) The coding sequence for SARS‐CoV‐2 proteins. The orf1ab encodes the pp1ab protein that contains 15 nsps (nsp1‐nsp10 and nsp12‐nsp16). The orf1a encodes the pp1a protein that contains ten nsps (nsp1‐nsp10). SARS‐CoV‐2 encodes four structural proteins spike (S), envelope (E), membrane (M), and nucleocapsid (N) and eight accessory proteins 3a, 3b, p6, 7a, 7b, 8b, 9b, and ORF14. 20 (b) Schematic of SARS‐CoV‐2 S protein. Similar to SARS‐CoV, S protein of SARS‐CoV‐2 also consists of (a) a signal peptide (SP; amino acids 1–19) located at the N terminus, (b) an extracellular domain (amino acids 20–1213) containing S1 (N‐terminal domain [NTD]: amino acids 20–286; C‐terminal domain [CTD]/RBD: amino acids 319‐541) and S2 (fusion peptide [FP] and Heptad repeat [HR1 and HR2]: amino acids 686–1213), (c) a transmembrane domain (TM: amino acids 1214–1236) and (d) a short cytoplasmic domain (CT: amino acids 1237–1273). 25 , 32 The D614G amino acid change in S protein is caused by an A‐to‐G nucleotide mutation at position 23,403 in the Wuhan reference strain. 13 The residue numbers of each region denote their positions in the S protein of SARS‐CoV‐2. CT, cytoplasmic domain; FP, fusion peptide; HR1/2, heptad repeat 1/2; NTD, N‐terminal domain; RBD, receptor‐binding domain; SARS‐CoV‐2, severe acute respiratory syndrome‐coronavirus‐2; SP, signal peptide; TM, transmembrane domain. The positions of N‐linked glycosylation sequons are shown as branches
Phylogenetic analysis of full‐length genomes indicated that SARS‐CoV‐2 is more closely related to bat‐SL‐CoV ZC45 and bat‐SL‐CoV ZXC21 (but more distantly related to SARS‐CoV) 11 and maximum homology (96.2% nucleotide sequence identity) with CoV RaTG13 isolated from Rhinolophus affinis bats. 21 Interestingly however, in phylogenetic analyses based on receptor‐binding domain (RBD) of the S or the S gene regions, SARS‐CoV‐2 was more closely related to SARS‐CoV, indicating the high sequence similarity of S gene between two viruses. 11 Accordingly, the origination of SARS‐CoV‐2 is commonly believed to be through the recombination of bat SARS‐CoVs with most frequent recombination breakpoints located within the ‘S’ gene. 22 To date, there are two propositions to explain the origin of SARS‐CoV‐2. The first scenario is based on high genomic sequence similarity (96%) between SARS‐CoV‐2 and the CoV isolated from a bat in 2013 (bat CoV RaTG13) and suggests a possible homologous recombination between the bat CoV and another CoV of unknown origin. 23 The second scenario is based on natural selection in humans following zoonotic transmission. 21 , 24 Indeed, S protein plays an essential role in viral attachment, fusion and entry into the host cells and might be the key protein for crossing the species barrier for adaptive evolution and animal‐to‐human transmission of SARS‐CoVs. 25 , 26 It is shown that nAbs targeting S protein block virus interaction with ACE2, while T‐cell responses against the SARS‐CoV‐2 S protein correlate with IgG and IgA antibody titres in Covid‐19 patients. 27 , 28 , 29 , 30 Therefore, the S protein has attracted particular attention as the most likely target antigen for long‐term immune response and vaccine design to SARS‐CoV‐2.
The present manuscript, reviews different characteristics of S protein, its potency and ‘state of the art’ of the vaccine development strategies and platforms using this antigen, for construction of a safe and effective SARS‐CoV‐2 vaccine.
2. STRUCTURAL/FUNCTIONAL FEATURES OF S PROTEIN
The S gene encodes a 1273 amino acid protein which is heavily glycosylated during its synthesis and assembles into trimers on the virion surface, resulting to the crown‐like appearance or corona. 31 Schematic diagram of the S protein and its various domains is presented in Figure 1b. Two functional subunits S1 and S2 that arise from proteolytic processing are responsible for binding to the host cell receptor and fusion, respectively. Although the S1 subunits of SARS‐CoV and SARS‐CoV‐2 can bind ACE2 to infect humans, the affinity of the RBD in the S1 subunit to ACE2 in SARS‐CoV‐2 is 10 to 20 fold stronger than that of the SARS‐CoV, which may contribute to the higher spread rate of SARS‐CoV‐2 from human to human. 32 Unlike SARS‐CoV, the S protein of SARS‐CoV‐2 contains a polybasic four residues at the boundary between the S1 and S2 subunits (a furin cleavage site) that might contribute to the tropism and transmissibility of SARS‐CoV‐2. 21
The cryo‐electron microscopy (Cryo‐EM) data of SARS‐CoV and MERS‐CoV S proteins indicated that the binding of S1 subunit to the host cell receptor forms a metastable pre‐fusion conformation (‘up’/‘opened’ conformation and /or ‘down’/‘closed’ conformation) that switches a stable post‐fusion conformation in the S2 subunit to facilitate the fusion steps. Such up and down conformations might be responsible for receptor‐accessible and receptor‐inaccessible states, respectively. Accordingly, recent studies on SARS‐CoV‐2 (Figure 2) indicated the presence of trimers with only a single RBD up protomer. 25 , 32 This finding suggests that unstable, distinct conformational states might lead to the initiation of fusogenic conformational change similar to the highly pathogenic CoVs (SARS‐CoV and MERS‐CoV). 33 , 34 This is in contrast to the common cold‐related CoVs that have RBD down conformation in the S trimers. 35 , 36 , 37 , 38 It should be noted however that in case of HCoV‐NL63 and HCoV‐229E with closed S trimers, RBDs hidden at the interface between protomers might need to be exposed. 39 , 40 Overall, these findings emphasize that S protein trimers in highly pathogenic CoVs seem to exist in partially opened (up) state, whereas they remain largely closed (down) in CoVs associated with the common cold.
FIGURE 2.
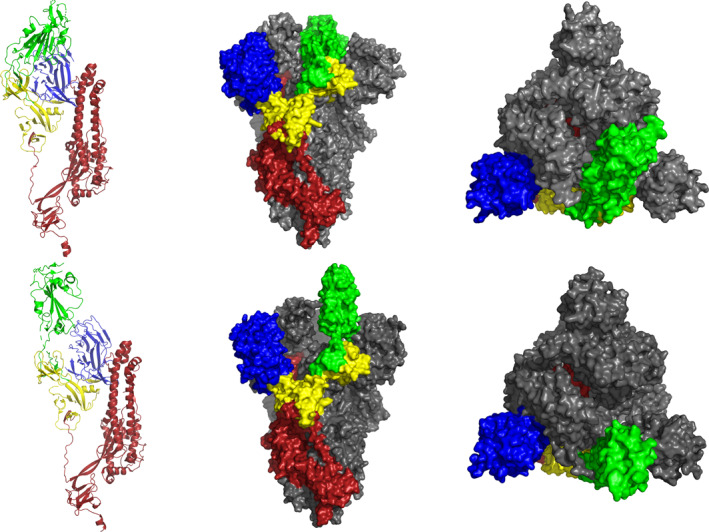
Structure of SARS‐CoV‐2 S protein in the pre‐fusion conformation. In the top row, the ribbon diagram shows single protomer of SARS‐CoV‐2 S consists the RBD (green) in the down conformation (closed RBD; left) and surface diagrams show side (centre) and apical (right) views of the structure of SARS‐CoV‐2 S trimer with three RBD (green and grey) in the down conformation (closed SARS‐CoV‐2 S trimer). In the bottom row, ribbon diagram shows single protomer of SARS‐CoV‐2 S consists the RBD (green) in the up conformation (opened RBD; left) and surface diagrams show side (centre) and apical (right) views of the structure of SARS‐CoV‐2 S trimer with single RBD (green) in the up conformation (opened SARS‐CoV‐2 S trimer). The structure of the SARS‐CoV‐2 S protein (PBD ID: 6VSB) 32 were analysed and modelled with molecular visualization software VMD (Version 1.9.3). RBD, receptor‐binding domain; SARS‐CoV‐2, severe acute respiratory syndrome‐coronavirus‐2
The SARS‐CoV‐2 S protein contains 22 N‐linked glycosylation sequons per protomer that contain oligomannose and complex glycans (Figure 1b). Glycosylation is critical to folding of S glycoprotein and immune evasion by shielding specific epitopes from antibody neutralization. Of note, several proximal glycosylation sites (N165, N234, N343) are able to mask RBD on S trimer, especially in RBD closed or down conformation. 25 , 41
3. POTENTIAL CORRELATES OF PROTECTIVE IMMUNITY TO SARS‐CoV‐2 AND THE ROLE OF S PROTEIN
Despite uncertainty about immunological correlates of protection for Covid‐19, correlation of virus‐specific nAbs titres and the numbers of virus‐specific T cells to SARS‐CoV‐2 (specially against S protein) with effective clearance of virus is reported in several studies (outlined in the following).
3.1. B cell immune responses and nAbs against SARS‐CoV‐2
It is well known that the humoral immune response is the critical primary effector of protective immunity for natural viral infection and vaccines. In case of Covid‐19, seroconversion in most of the infected people occur between 7 and 14 days after the onset of symptoms, starting with the detection of IgM and IgA antibodies (that can be detected early during the first week or 3 weeks of symptom onset) followed by IgG detection by around 14 days after the initiation of symptoms 28 , 42 , 43 (Figure 3). Rise in the Ab levels is also accompanied by the increase in activated CD4+/CD8+ T‐cells and plasma cells in peripheral blood mononuclear cells (PBMCs) 44 , 45 while IgG memory cells specific to the RBD have also been detected in the blood of Covid‐19 patients. 46 Similarly, the prevention of reinfection in SARS‐CoV‐2 infected rhesus macaques correlated with the rise of antibodies in recovered animals. 47 In parallel, several studies in infected patients have shown the presence of serum IgA against SARS‐CoV‐2 with neutralizing potential 42 , 48 as shown previously in preclinical animals studies (in bronchoalveolar lavages) with SARS‐CoV vaccine candidates. 49 , 50
FIGURE 3.

Immune response to SARS‐CoV‐2. Dendritic cells as APC present viral antigens to CD4+ T cells and induce the production of IgG, IgM and IgA to prevent viral entrance. Furthermore, cytokine storm starts in severe cases that might be correlated with disease severity. It was shown that antibodies and CD4+ T cells generated in 100% of recovering Covid‐19 patients. The CD8+T cells also detected in 70% of recovering COVID patients which secrete perforin and granzyme to kill virally infected cells. It was found CD4+ T‐cell responses to S protein, the main target of most vaccine efforts, were robust, and correlated with the magnitude of the anti‐SARS‐CoV‐2 IgG and IgA titres. T cell responses are focused not only on S but also on M, N, and other ORFs. APC, antigen‐presenting cell; Covid‐19, coronavirus disease 2019; SARS‐CoV‐2, severe acute respiratory syndrome‐coronavirus‐2
In general antibodies against both the N and S proteins are commonly detectable, among which those raised against RBD of S protein can be potently neutralizing and could be detected in most tested Covid‐19 patients. 28 , 46 , 51 Of note, neither plasma of convalescent Covid‐19 patients nor SARS‐CoV‐2 RBD‐specific neutralizing monoclonal antibodies (mAbs) showed any cross‐reactivity with that of the SARS‐CoV or MERS‐CoV. However, that of the SARS‐CoV showed cross‐reactive neutralization with SARS‐CoV‐2 26 , 52 , 53 , 54 indicating the possibility of using SARS‐CoV S (RBD) as the antigen to induce nAbs against SARS‐CoV‐2. Indeed, several mAbs and nanobodies derived against the S1‐RBD, S1‐NTD, and S2 of SARS‐CoV and MERS‐CoV might confer cross‐activity against virus SARS‐CoV‐2 viral entry. 27 , 52 , 55 , 56 , 57 , 58 , 59 It was reported that that SARS‐CoV specific human mAbs, s309 60 and CR3022, 52 , 54 were capable of binding to the SARS‐CoV‐2 effectively. 61 Accordingly, sera from recovered patients of Covid‐19 (as a potential source of nAbs) were used to generate mAbs against SARS‐CoV‐2. Four of the generated mAbs (31B5, 32D4, P2C‐2F6 and P2C‐1F11) indicated high neutralizing activity in vitro by efficiently inhibiting ACE2‐RBD binding. 25 , 46 , 62 , 63 Alternatively, mAbs 47D11 and n3130 produced from SARS‐CoV and SARS‐CoV‐2 respectively were shown to neutralize SARS‐CoV‐2 without inhibiting ACE2‐RBD binding. 64 , 65 In several other recent studies, Abs from convalescent Covid‐19 patients (which are correlated with the S1, RBD and S2 regions) were used to treat SARS‐CoV‐2 infection. 27 , 28 Animal models were also used to generate nAbs against SARS‐CoV‐2. In this regard, nanobodies, containing a variable heavy (VH) chain against SARS‐CoV‐2 66 or SARS‐CoV and MERS CoV S proteins fused to a human Fc fragment (VHH‐72‐Fc), were developed in immunized camelids. 66 Besides, construction of recombinant ACE2‐Fc and RBD‐Fc fragments 67 , 68 with in vitro cross‐neutralizing activity for both SARS‐CoV and SARS‐CoV‐2 was also reported.
It should be noted that despite strong evidence on the correlation of anti‐SARS‐Cov‐2 antibody titres to higher rates of viral neutralization (in vitro) and decline of viral load in patients (in vivo), 63 , 69 , 70 some severe clinical cases of Covid‐19 persisted despite the presence of higher antibody titres. 69 , 70 , 71 This scenario which was also reported in the previous SARS‐CoV and MERS‐CoV epidemics, 72 , 73 , 74 , 75 raised concerns about antibody‐dependent enhancement (ADE). This phenomenon occurs when non‐nAbs against proteins of a virus enhance virus entry to host cells particularly macrophages and monocytes, also enhancing virus infectivity and inflammatory activation. 76 It should be noted that to date, there is no report on contribution of anti‐SARS‐CoV‐2 antibodies to the pathological features observed in Covid‐19 patients.
Taken together, from findings of resent studies, it might be concluded that S1 and particularly RBD could be considered as the main antigen candidates in vaccine platform formulations to induce virus‐specific nAbs to prevent SARS‐CoV‐2 infection.
3.2. T cell response
Although specific antibody responses are the primary effector of protective immunity against viral infections, T cell responses appear to play vital roles in the clearance of several viruses. 77 Concerning Covid‐19 infection, occurrence of lymphocytopenia (decline in the lymphocyte count) in both CD4 and CD8 T cells and decreased levels of circulating B cells, natural killer (NK) monocytes, eosinophils and basophils in severe cases is shown. 14 , 78 , 79 , 80 Moreover, most of the severe Covid‐19 cases (especially in ICU patients) exhibited significantly increased serum levels of pro‐inflammatory cytokines and chemokine (so‐called ‘cytokine storm’: e.g., IL‐6, IL‐1β, IL‐2, IL‐8, IL‐17, G‐CSF, GMCSF, IP‐10, MCP‐1, CCL3 and TNFα) which correlated with the reduced number of T cells and severity of the disease. 78 , 79 , 81 In Covid‐19‐induced severe pneumonia, higher levels of nearly similar cytokines/chemokines were correlated to lung injury, indicating that the cytokine storm and exacerbated inflammatory responses were manifested clinically by acute respiratory distress syndrome (ARDS) 82 (Figure 3). Such cytokine storms (or cytokine‐release reactions), represent a hypersensitivity reaction (HSR) via activation of various immune cells by HSR mediators (i.e.,: IL‐6). 83 Moreover, in cytokine storm reactions, complement component C3a and C5a bind to complement receptors resulting in the release of histamine, leukotrienes, and prostaglandins and contribute to the main symptoms such as flushing, hives, hypoxia, vasodilation and hypotension. Indeed, the representative SARS‐CoV ssRNAs have powerful immuno‐stimulatory activities in releasing pro‐inflammatory cytokines (TNF‐α, IL‐6 and IL‐12). 83 Elevated levels of some pro‐inflammatory cytokines (MCP‐1, TGF‐ β1, TNF‐α, IL‐1 and IL‐6) produced by SARS‐CoV infected cells, might cause acute lung injury (ALI). Accordingly, in H5N1 influenza A viral infection, the inflammatory cytokines such as IL‐1β, IL‐8 and IL‐6 play a major role in mediating and amplifying ALI and ARDS by stimulating C5a chemotaxis. The C5a induces innate immune cells (mast cells, neutrophils and monocytes/macrophages) to release pro‐inflammatory cytokines such as IL‐12, TNF‐α and macrophage inflammatory proteins‐1α. In addition, C5a also stimulates adaptive immune cells such as T and B cells to release cytokines such as TNF‐α, IL‐1β, IL‐6 and IL‐8. Similar studies have also shown that H7N9‐infected patients have significantly higher levels of cytokines such as IL‐6, IP‐10, IL‐10, IFN‐γ and TNF‐α compared to healthy volunteers. These observations indicate that the cytokine storm reaction might play an important role in ALI. 83
Among main SARS‐CoV proteins (S, N and M, as well as ORF3), T cell responses against the S and N proteins have been documented to be the most dominant and long‐lasting. 84 , 85 Despite the short‐lived antibody responses in convalescent Covid‐19 patients, T cell responses were shown to induce long‐term protection. 86 , 87 , 88 Several predicted T cell epitopes within the S protein of SARS‐CoV are completely identical to SARS‐CoV‐2 89 , 90 implying the potential to elicit cross‐protection responses. Recently, validation of the predicted T cell epitopes are undertaken by MegaPools, using PBMCs from patients recovered from Covid‐19 disease as well as unexposed individuals. 20 Results indicated that specific CD8+ and CD4+ T cells were generated in around 70% and 100% of Covid‐19 patients, respectively. 29 The identified CD4+ T cell responses were strong and associated with the induction of IgG and IgA antibody. Of note, 50% of the total CD4+ T cell responses were against the S protein, while the specific CD8+ T cells against S protein were also found in most, if not all participants. 29 , 91 In another study, 13 out of 14 recovered Covid‐19 patients, showed strong correlation between nAb titres and the numbers of virus‐specific T cells. 92 Accordingly, results of a study on ten Covid‐19 patients with moderate to severe ARDS showed strong and specific CD4+ and CD8+ cells mostly against S protein in 100% and 80% of patients, respectively. 93 These cellular responses were mainly skewed towards Th1, although Th2 and Th17 cytokines were also found. Besides, low levels of specific T‐cells were found in 20% of unexposed individuals as a potential indicator of cross‐reactive T cell between SARS‐CoV‐2 and common cold‐causing coronaviruses. 93 In parallel, results of a cohort study indicated SARS‐CoV‐2‐specific CD4+ and CD8+ T cells with high cytotoxic activity in the acute phase of the disease 94 (specially S protein specific) implying the role of cellular response in a potential vaccine.
4. VACCINE CANDIDATES BASED ON SPIKE (S) PROTEIN
Results of prior animal studies with first generation (whole virus‐based inactivated and attenuated vaccine modalities) SARS‐CoV 76 , 95 , 96 or MERS‐CoV vaccines 75 , 97 , 98 , 99 indicated the possibility of adverse effects such as increased infectivity and immuno‐potentiation (in the form of eosinophilic infiltration) and/or ADE in immunized animals. Considering that both SARS‐CoV‐2 and SARS‐CoV share a high degree of genetic and pathologic identity, it is reasonable to think that a whole virus‐based SARS‐CoV‐2 vaccine might also induce the same adverse effects. Therefore, development of second and third generation safer vaccines (by using modern vaccine platforms) using a proper viral protein as vaccine antigen is actively sought for SARS‐CoV‐2 infection.
Based on information provided for S protein (being the main determinant of cell entry and tropism, containing protective B‐ and T‐cell epitopes), 29 , 100 this SARS‐CoV‐2 surface protein might be an ideal candidate antigen for modern vaccine platforms and modals to produce a safe vaccine (Figure 4). To this end, various platforms, including viral vectors (replicating and non‐replicating), nucleic acids (DNA and RNA), recombinant proteins and virus‐like particles in various formulation strategies including polymer‐ and lipid‐based nanoparticles (for nucleic acid encapsulation), and adjuvants based on aluminium or saponin as well as Toll‐like receptor (TLR) agonists have been investigated to elicit potent immune responses against either full‐length or fractions of the SARS‐CoV‐2 S glycoprotein. 101 , 102
FIGURE 4.
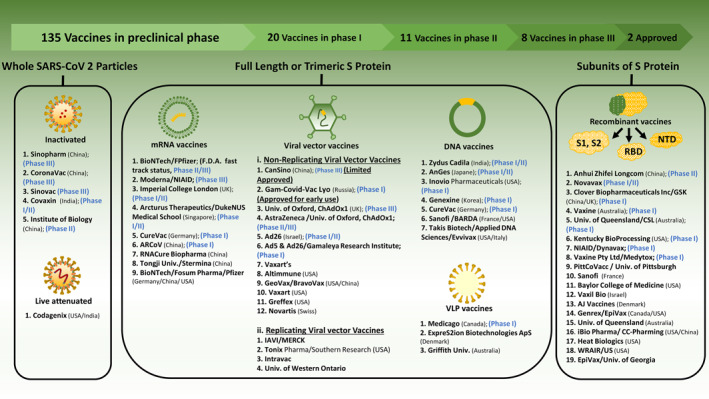
Potential and developing candidates of SARS‐CoV‐2 vaccine platforms. SARS‐CoV‐2, severe acute respiratory syndrome‐coronavirus‐2
It should be noted that, despite reports on safety of SARS‐(full) S protein‐encoding vaccines in immunized mice or non‐human primates 103 , 104 , 105 , 106 and mice immunized passively by anti‐S‐antibody, 107 , 108 but ADE has been observed in cats vaccinated by recombinant vaccinia virus expressing fusogenic S protein. 109 In addition, lung immunopathology and hepatitis have been found in SARS‐CoV‐challenged animal models after vaccination with SARS‐(full) S protein‐encoding vaccines, the same as that of whole viral‐vaccine. 95 , 96 , 110 , 111 These observations resulted to application of various segments of SARS‐CoV‐2 S protein including RBD, NTD, S1 and S2 (besides full S fragment). It is reported that the S1 subunit or RBD of S protein induce nAbs without potential of ADE development. 76 , 112
4.1. The full‐length S protein based vaccines
In several previous studies, the full‐length S protein was used to develop SARS‐CoV and MERS‐CoV vaccine candidates. DNA vaccines encoding S protein of SARS‐CoV Urbani strain was shown to induce immune responses that protected mice model against virus challenge. 103 , 113 Moreover, the DNA vaccine encoding MERS‐CoV S protein was effective in eliciting both nAbs and cellular immune responses that protected immunized non‐human primates against virus challenge. 104 In parallel, several animal immunization studies using viral platforms expressing full‐length S protein of the SARS‐CoV, reported promising protective results against viral infection. Highly attenuated modified vaccinia virus Ankara (MVA) induced nAb against S protein and decreased virus shedding in the respiratory tracts of mice or monkeys after virus challenge. 105 , 106 Likewise, a recombinant attenuated parainfluenza virus expressing the full length S protein of SARS‐CoV protected immunized monkeys from subsequent homologous SARS‐CoV challenge. 107 Accordingly, the administration of full‐length S protein trimer to mice or hamsters was also shown to induce significant protection against homologous virus shedding. 108 Similarly, animal immunization with baculovirus, expressing the full‐length and extracellular domain of S protein from the SARS‐CoV Urbani strain was shown to induce nAbs against homologous and heterologous pseudoviruses of SARS‐CoV. 114 Recently, it was shown that SARS‐CoV and MERS‐CoV S nanoparticles produced in the baculovirus expression system induce high titres of nAbs against the homologous but not the heterologous virus (i.e., no cross‐protection). 115
Currently, several developers use full‐length S protein as antigen in various platforms to construct an efficient vaccine candidate against SARS‐CoV‐2 that are currently in the clinical trial or preclinical phases (Table 1: based on WHO draft landscape of Covid‐19 candidate vaccines–28 September 2020). 116 Four well‐known types of such vaccines are ChAdOx1 nCoV‐19 (University of Oxford/AstraZeneca), Ad5 vector (CanSino Biological Inc./Beijing Institute of Biotechnology), mRNA‐1273 (Moderna/NIAID) and BNT162b2 (BioNTech/Fosun Pharma/Pfizer), 117 , 118 , 119 , 120 , 121 and NVX‐CoV2373 recombinant protein (Novavax) 122 that based on promising results on induction of protective nAbs in animal models have entered phase I clinical trials and now are undergoing phase III evaluation.
TABLE 1.
Vaccine types and platforms with full‐length S protein (disclosed) as antigen formulation that are engaged in clinical stages (based on data from 28 September 2020 update 116 )
| Platform | Vaccine specifications | Developer | Phase of clinical evaluation |
|---|---|---|---|
| Non‐replicating viral vector | Chimpanzee adenovirus | University of Oxford/AstraZeneca | Phase 3 |
| Ad5 | Cansino Biological Inc./Beijing Institute of Biotechnology | Phase 3 | |
| Ad26 | Janssen Pharmaceutical Companies | Phase 3 | |
| Ad5 and Ad26 | Gamaleya Research Institute | Phase 3 | |
| Simian | ReiThera/LEUKOCARE/Univercells | Phase 1 | |
| Adenovirus | |||
| Ad5 | Institute of Biotechnology, Academy of Military Medical Sciences, PLA of China | Phase 1 | |
| Ad5 (oral vaccine platform) | Vaxart | Phase 1 | |
| Replicating viral vector | Measles‐vector based | Institute Pasteur/Themis/Univ. of Pittsburg CVR/Merck Sharp & Dohme | Phase 1 |
| mRNA | LNP‐encapsulated | Moderna/NIAID | Phase 3 |
| LNP‐encapsulated | BioNTech/Fosun Pharma/Pfizer | Phase 3 | |
| LNP‐encapsulated | Curevac | Phase 2 | |
| SAM | LNP‐encapsulated | Arcturus/Duke‐NUS | Phase 1/2 |
| Based on VEEV vector | Imperial College London | Phase 1 | |
| DNA | Plasmid DNA with electroporation | Inovio Pharmaceuticals | Phase 1/2 |
| Plasmid DNA | Genexine Consortium | Phase 1/2 | |
| Protein subunit | Adjuvanted with Matrix M | Novavax | Phase 3 |
| S‐trimer adjuvanted with MF59 | University of Queensland/CSL/Seqir | Phase 1 | |
| S‐trimer adjuvanted with AS03 and CpG 1018 | Clover Biopharmaceuticals Inc./GSK/Dynavax | Phase 1 | |
| S‐2P adjuvanted with CpG 1018 | Medigen Vaccine Biologics | Phase 1 | |
| Corporation/NIAID/Dynavax | |||
| Adjuvanted with Advax™ | Vaxine Pty Ltd/Medytox | Phase 1 |
Abbreviations: Ad5, human adenovirus type 5; Ad26, human adenovirus type 26; LNP, lipid nanoparticle; SAM, self‐amplifying mRNA; VEEV, Venezuelan equine encephalitis virus.
The ChAdOx1 nCoV‐19 vaccine (AZD1222) is a replication‐deficient simian adenovirus vector, containing the full‐length codon‐optimized coding sequence of SARS‐CoV‐2 S protein along with a tissue plasminogen activator (tPA) leader sequence. Preclinical immunogenicity of the ChAdOx1 nCoV‐19 was assessed in two mouse strains (BALB/c and outbred CD1) and rhesus macaques. Intramuscular (IM) injection of 6 × 109 virus particles (VPs) in mice induced detectable total IgG titres and virus‐specific nAbs in all vaccinated mice and IgG subclass profiling showed a predominantly Th1 response. The Th1‐type response was also supported by high levels of IFN‐γ and TNF‐α, and low levels of IL‐4 and IL‐10 post‐vaccination. In rhesus macaques, virus‐specific nAbs and T‐cell responses were induced in all tested animals 14 days after IM injection of 2.5 × 1010 VPs. Vaccinated macaques that were challenged with SARS‐CoV‐2 had significantly reduced viral load in bronchoalveolar lavage fluid and respiratory tract tissue, and showed no pneumonia and evidence of immune‐enhanced disease following viral challenge. 118 Phase 1/2 clinical trial of ChAdOx1 nCoV‐19 was performed by a single‐dose or two‐dose IM injection of 5 × 101⁰ VPs. Humoral responses to S protein maximized by Day 28 post first dose, and cellular responses were induced in all participants by Day 14. After two doses, nAbs and potent cellular and humoral immune responses were induced in all participants. The vaccine was safe and well‐tolerated, and no serious adverse events were noted. 117
The Ad5 vector vaccine developed by CanSino Biological Inc is an E1 and E3 deleted Ad5 vector containing an optimized full‐length S protein of SARS‐CoV‐2, with the tPA signal peptide. In phase 1 clinical trial, three doses including low dose (5 × 1010 VPs), middle dose (1 × 1011 VPs) and high dose (1.5 × 1011 VPs) were given to different groups of participants as a single IM injection. Humoral responses peaked at Day 28, and specific T‐cell responses were detected from Day 14 post vaccination in healthy adults. In phase 2 trial, single doses of 1 × 1011 VPs or 5 × 1010 VPs were similarly administered IM. Both doses of the vaccine induced significant nAb responses. The vaccine induced immune responses within 14 days, and 95% of participants receiving 1 × 1011 VPs and 91% of the recipients receiving 5 × 101⁰ VPS showed either cellular or humoral immune responses at Day 28 post‐vaccination. Increased IFN‐γ‐producing T‐cells were found in 90%, and 88% of participants receiving 1 × 1011 and 5 × 1010 VPs, respectively. While no serious adverse reactions were reported, some adverse events were documented in 9% of participants in the 1 × 1011 VPs dose group and 1% participant in the 5 × 1010 VPs dose group. Accordingly, the Ad5 vector‐based vaccine was considered to be safe at the dose of 5 × 1010 VPs and was capable of inducing considerable immune responses in the majority of recipients after a single immunization. Despite presence of high and low pre‐existing anti‐Ad5 nAb in 52% and 48% of participants, respectively (as one shortcoming of the vaccine) that along with increasing age could partially hinder the humoral immune responses, 120 the vaccine has recently received military specially needed drug approval in China.
Moderna's LNP‐encapsulated mRNA vaccine candidate (mRNA‐1273) encodes the pre‐fusion conformation of S glycoprotein with a transmembrane anchor that has been stabilized by two consecutive proline substitutions (S‐2P) of residues 986 and 987. In preclinical evaluation, BALB/c, C57BL/6 and B6C3F1 mice strains were immunized by two‐dose IM injection of 0.01, 0.1 or 1 μg of mRNA‐1273. The antibody titres increased with dose level, and a potent neutralizing activity was induced by 1 μg of the vaccine, while the 10 μg dose elicited robust neutralizing activity. The IgG and cytokine profiles demonstrated that immunization with mRNA‐1273 induced a balanced Th1/Th2 response. 120 In phase 1 clinical trial, 25, 100 or 250 μg of the vaccine was injected as either single‐dose or two‐dose intramuscularly. After the first vaccination, antibody responses were higher with the higher dose, and after the second vaccination, the titres increased, and serum neutralizing activity was detected in all participants. The Th1‐skewed CD4 T cell response supported by high expression of TNFα, IL‐2 and IFN‐γ, and minimal expression of IL‐4 and IL‐13, was detected in 25 and 100 μg dose groups. The 100 μg dose elicited low levels of specific CD8 T cell responses after the second vaccination. Among the three doses, the 100‐μg dose induced high neutralization responses and Th1‐biased CD4 T cell responses, along with a safety profile that was more favourable than that of the higher dose. No serious adverse events were documented, but systemic adverse reactions were more common after the second vaccination, predominantly with the 250 μg dose. In the phase 2 trial, each participant has been allocated to receive a double IM injection of 50 μg or 100 μg doses. 123
BNT162b2 is a 1‐methylpseudouridine (m1Ψ)‐modified mRNA encapsulated in LNP (m1Ψ–mRNA‐LNPs) that encodes S‐2P mutant form of the full‐length S protein. In preclinical evaluation, BALB/c mice were injected intramuscularly once with 0.2, 1 or 5 µg of the antigen, and rhesus macaques (Macaca mulatta) were immunized by two‐dose IM injection of 30 or 100 µg of BNT162b2 on Days 0 and 21. A single injection of BNT162b2 in mice elicited high neutralizing titres and strong Th1 and Tfh type CD4+ and IFNγ+ IL‐2+ CD8+ T‐cell responses. The immunogenicity of BNT162b2 in rhesus macaques was parallel to the immunogenicity in mice. The rhesus macaques that had received two immunizations with 100 µg BNT162b2 were challenged with 1.05 × 106 plaque‐forming units of SARS‐CoV‐2 USA‐WA1/2020 isolate that was performed 55 days after the second immunization. The BNT162b2 vaccine candidate fully protected the lungs of immunized rhesus macaques from the SARS‐CoV‐2 challenge. 124 In phase I clinical trial, 10, 20 or 30 μg of the vaccine was injected intramuscularly as two‐dose, 21 days apart. Data on immune responses or safety beyond 7 days after the second dose were not available until publication date of the report, although at 7 days after the second dose, the SARS‐CoV‐2–neutralizing geometric mean titres (GMT), elicited by 30 µg BNT162b2, was significantly exceeded the GMT of the convalescent serum panel. The trial has now gotten advanced at the 30‐μg dose level into the phase 2/3. 121
Novavax's NVX‐CoV2373 is based on the codon‐optimized S‐2P mutant form of full‐length S‐protein with an additional mutation of the furin cleavage site (682‐RRAR‐685) to 682‐QQAQ‐685. The protein is produced in the baculovirus‐Spodoptera frugiperda (Sf9) insect cell expression system and is adjuvanted with the saponin‐based Matrix‐M1. For preclinical evaluation, BALB/c mice were immunized by IM injection with a single dose or two doses spaced 14 days apart containing a dose range (0.01, 0.1, 1.0 or 10 μg) of NVX‐CoV2373 with 5 μg saponin‐based Matrix‐M1 adjuvant. Also, the olive baboons (Papio cynocephalus anubis) were immunized twice (21 days apart) by IM injection with 1, 5 or 25 μg NVX‐CoV2373 with 50 μg Matrix‐M1 adjuvant. For virus challenge in mice, the mice were transduced intranasally with 2.5 × 108 pfu Ad/CMVhACE2 38 days after the second vaccination. At 4‐day post‐infection, the mice were intranasally inoculated with 1.5 × 105 pfu of SARS‐CoV‐2. Immunizations of mice and baboons elicited multifunctional CD4+ and CD8+ T cell responses with a Th1 biased phenotype, along with SARS‐CoV‐2 neutralizing antibodies. In the challenged mice, the vaccine was protective with no indication of vaccine‐associated enhanced respiratory disease. 125 The preclinical evaluation was promoted by immunizing cynomolgus macaques (Macaca fascicularis) with two‐dose (21 days apart) IM injection of 5 or 25 μg NVX‐CoV2327 with 50 μg Matrix‐M1. Animals were challenged with 1.04 × 104 pfu SARS‐CoV‐2 isolate USA‐WA1/2020 intranasally and intratracheally. The vaccine administration induced anti‐S neutralizing antibody and protected macaques against upper and lower infection and pulmonary disease. 126 In phase I/II trial, the participants received two IM injections of either 5 or 25 μg of NVX‐CoV2373 plus Matrix‐M1 on Days 0 and 21. At 35 days, no serious adverse events were documented, and a Th1‐biased response was elicited along with SARS‐CoV‐2 neutralizing GMT levels approximately four times greater than those in symptomatic outpatients with Covid‐19. 122
4.2. The RBD of S protein based vaccines
Several previous studies on SARS‐CoV and MERS‐CoV showed presence of B‐cell epitopes in RBD of S proteins capable of inducing nAbs to block the interaction of RBD with cellular receptor. 127 Accordingly, from 27 developed mAbs against SARS‐CoV RBD, 23 showed neutralizing activity, 54 , 114 among which some bind to RBM within the RBD, while others bind to domains outside this region within RBD. 54 , 114 Of note, a number of prior studies reported the presence of several conformational B‐cell epitopes in recombinant RBDs, capable of inducing of cross‐reactive nAbs against SARS‐CoV and MERS‐CoV. 128 , 129 , 130 , 131 , 132 Accordingly, the high reactivity of such nAbs towards SARS‐CoV pseudoviruses was reported. 133 In addition, several studies reported the strong interaction of RBD with nAbs in the antisera of either patients infected with SARS‐CoV (in the convalescent phase) or animal models immunized by full‐length S protein expressing‐MVA. 106 , 134 Furthermore, rabbits immunized by a RBD‐Fc human IgG fusion protein could generate a potent neutralizing activity and long‐term protection against homologous SARS‐CoV challenge. 135 , 136 Similarly, immunization with the RBD‐Fc fusion protein elicited nAbs to protect hCD26/DPP4 transgenic mice against MERS‐CoV infection. 137 Mice models immunized by vector‐based vaccines (such as an adeno‐associated virus‐expressing RBD) developed nAbs that protected the animals from homologous virus challenge. 138 , 139 , 140 It should be noted however that while induction of nAbs against RBD is the primary effector response of the protective immunity, T‐cell immune responses that might further contribute to the protection were also found following immunization of mice with the RBD‐based subunit vaccines. 33 , 140 , 141 Currently, there are two LNP/mRNA platform (BioNTech/Fosun Pharma/Pfizer [phase 1/2], and People's Liberation Army [PLA] Academy of Military Sciences/Walvax Biotech [phase 1]), and six protein subunit platforms (Anhui Zhifei Longcom Biopharmaceutical/Institute of Microbiology, Chinese Academy of Sciences [phase 2] and Kentucky Bioprocessing Inc [phase 1/2], Instituto Finlay de Vacunas of Cuba [phase 1], West China Hospital of Sichuan University [phase 1], COVAXX [phase 1] and an RBD‐HBsAg VLP by SpyBiotech/Serum Institute of India [phase 1/2]), and one flu‐based replicating viral vector (Beijing Wantai Biological Pharmacy/Xiamen University (phase 1)) in the clinical trial that use RBD of SARS‐CoV‐2 as the vaccine candidate. 116 BioNTech's BNT162b1 LNP‐encapsulated mRNA vaccine candidate encodes the RBD of the SARS‐CoV‐2 S protein, linked to a foldon trimerization domain to increase its immunogenicity through repetitive antigenic display. The RNA is optimized by incorporating 1‐methyl‐pseudouridine instead of uridine to reduce innate immune sensing and to increase in vivo translation of mRNA. 142 Phase 1/2 trial of the vaccine has been performed in Germany 142 and the USA. 143 Two doses of 1–50 µg of the vaccine applied in Germany elicited potent CD8+ and Th1‐type CD4+ T cell responses, supported by high secretion of IFN‐γ. The sera from two injections demonstrated robust antibody responses with strong neutralization activity. 142 In the case of 10 and 30 µg dose levels applied in the US trial, IgG concentrations and neutralizing titres in sera increased with the dose level after a second dose. 143
There are still other platforms in the preclinical stage using RBD as the main antigen for SARS‐CoV‐2 vaccine development that include (i) the protein subunit vaccine developed by Baylor College of Medicine, Biological E Ltd, Mynvax, Chulalongkorn University/GPO Thailand, Neovii/Tel Aviv University, and Baiya Phytopharm/Chula Vaccine 116 ; (ii) the LNP‐encapsulated mRNA vaccine co‐developed by Fudan University/Shanghai JiaoTong University/RNACure Biopharma 116 ; (iii) plasmid DNA developed by Scancell/University of Nottingham/Nottingham Trent University, and National Research Centre of Egypt; (iv) the virus‐like particle (VLP) vaccine developed by Saiba GmbH 116 and (v) the replicating influenza vector vaccine developed by university of Hong Kong. 116
Taken together, it seems that candidate vaccines based on RBD of SARS‐CoV‐2 are supposed to have bright future and more attraction in near future.
4.3. The NTD of S protein based vaccines
Alike RBD, NTD in S protein of some CoVs show receptor‐binding activity through binding to sugar moieties. 144 , 145 Several studies showed that recombinant NTD protein of MERS‐CoV is capable of eliciting sufficient nAbs and cellular immune responses to protect against virus challenge in animal models. 33 , 146 , 147 Although compared to other regions of S protein (full‐length S protein, S1 and RBD), NTD is less immunogenic (i.e.,: eliciting considerably lower antibody titres and cellular immune responses), it might be involved in the binding of specific receptors 144 , 145 and thus deserve to be considered as a candidate antigen for vaccine development against Covid‐19.
4.4. The S1 subunit of S protein based vaccines
The S1 subunit, which contains both RBD and NTD regions, is responsible for virus binding to the host cell receptor. Prior studies indicated that the S1 subunit can induce strong immune responses and/or protection against viral infection. 58 , 148 Immunization of rats via subcutaneous or intranasal routes with a recombinant adenovirus encoding first 490 amino acids of the S1 subunit, elicited strong humoral immune responses that protected the animals against SARS‐CoV infection. 149 Similarly, immunization of hDPP4 transgenic mice with MERS‐CoV recombinant S1 protein, formulated with MF59 adjuvant, induced nAbs which correlated with protection. 150 A similar study also reported that intramuscular injection of an adjuvant formulated MERS‐CoV S1 protein (subunit vaccine) was capable of reducing virus shedding in dromedary camels, while conferring complete protection against the viral challenge in alpaca. 148 Recently, it was shown that subcutaneously immunized mice (either traditional needle injection or intracutaneously by dissolving microneedle arrays [MNAs]), by a codon‐optimized S1 subunit containing integrated (in‐built) TLR agonist sequences, elicited specific humoral responses which were of higher titres in MNAs delivery. 151 Therefore, the S1 subunit of SARS‐CoVs might also have the potential to be considered as the main antigen in different platforms to formulate a vaccine candidate against these viral infections.
To date, the vaccine candidates that use SARS‐CoV‐2 S1 as the primary antigen are in the preclinical stage and include a protein subunit vaccine platform co‐developed by AnyGo Technology (recombinant S1‐Fc fusion protein), University of Pittsburgh (microneedle arrays S1 subunit), and Baylor College of Medicine and also a recombinant deactivated rabies virus platform developed by Bharat Biotech/Thomas Jefferson University. 116
4.5. The S2 subunit of S protein based vaccines
The S2 subunit, which contains an internal membrane fusion peptide (FP) and heptad repeats (HR1 and HR2), is responsible for fusion between the viral and host cell membranes. The S2 subunit which is highly conserved among SARS‐CoVs and MERS‐CoV is an immunogenic protein. 89 , 152 , 153 , 154 , 155 Several studies have reported that HR1 and HR2 domains of S2 can generate broadly nAbs against pseudo‐typed heterologous SARS‐CoVs in vitro. 153 , 156 , 157 It should be noted however that other regions of S2 domain (residues 681–980) might elicit non‐nAbs (as shown in immunized mice). 158 In addition, an S2 peptide sequence (residues 736–761) of MERS‐CoV induced S2‐specific nAbs in rabbits, 131 although the protective efficacy is yet to be addressed. Recently, linear epitopes of SARS‐CoV‐2 S2 were mapped and the presence of cross‐reactive neutralizing epitopes for other SARS‐CoVs were shown, which sounds a more promising role for this protein as main antigen of a vaccine platform. 89 It should be noted however that the FP domain of S2 which is involved in the host cell membrane fusion and viral pathogenicity has also the potential of being used as an antigen of vaccine platforms, either alone or fused with other antigenic fragments (RBD, NTD, HR1 and HR2). To date, a RBD‐FP fusion protein that induced strong antibody response in immunized mice was constructed 159 but its protective efficacy remains to be addressed.
5. CONCLUSIONS
The ACE2 cellular receptor binding, S protein of SARS‐CoV‐2, plays an essential role in viral entry and infection and contains multiple B‐ and T‐cell epitopes to induce nAbs and long‐term protection, suggesting a main candidature for this protein as Vaccine antigen. Accordingly, three vaccines based on full‐length‐S antigen including two Ad‐based (ChAdOx1 nCoV‐19 and Ad5) and one RNA‐based (mRNA‐1273; Moderna/NIAID) were evaluated in phase I/2 clinical trials. With promising results on induction of nAbs and T‐cell immunity in immunized volunteers, these vaccines are now in the third clinical trial phase. Reports on some adverse effects (ADE development) on immunization with SARS‐(full) S protein‐encoding vaccines resulted to the application of various segments of SARS‐CoV‐2 S protein including RBD, NTD, S1 and S2 (besides full S fragment) in combination with various modern vaccine platforms. The RBD of SARS viruses, with potential of producing cross‐nAbs has been in the centre of concerns and currently two LNP/mRNA and two protein (subunit) based vaccines encoding RBD of SARS‐CoV‐2 are in phase 1/2 clinical trials, while several others are in preclinical studies. Results of 1/2 clinical trial on LNP/mRNA vaccines indicated induction of potent nAbs and cellular responses, implying the potential of early phase three studies. Induction of productive nAbs and cellular responses in preclinical studies on vaccine candidates encoding S1, S2 and NTD regions of S protein indicates potential of phase 1 clinical trials on such formulations in near future.
CONFLICT OF INTEREST
The authors declare no conflicting financial or other interests.
AUTHOR CONTRIBUTION
All authors contributed to the writing of the manuscript.
ACKNOWLEDGEMENTS
The members of the Rapid Response Team of PII also include Fatemeh Fotouhi, Ehsan Mostafavi, Monireh Kazemimanesh, Saber Esmaeili, Ali Maleki, Mahdi Rohani, Ahmad Ghasemi and Kazem Baesi. The authors are thankful to the departments of Molecular Virology, Arboviruses and Viral Hemorrhagic Fevers, Epidemiology, and ‘influenza & other Respiratory Viruses’ of PII for diagnostic and managerial affairs during SARS‐CoV‐2 outbreak.
Arash Arashkia and Somayeh Jalilvand have contributed to this work equally.
The members of Rapid Response Team of PII are listed in the Acknowledgements.
DATA AVAILABILITY STATEMENT
Data openly available in a public repository that issues datasets with DOIs.
REFERENCES
- 1. Chen Y, Liu Q, Guo D. Emerging coronaviruses: genome structure, replication, and pathogenesis. J Med Virol. 2020;92(4):418‐423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. de Wit E, van Doremalen N, Falzarano D, Munster VJ. SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol. 2016;14(8):523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Zhu N, Zhang D, Wang W, et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl J Med. 2020;382:727–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Peiris J, Guan Y, Yuen K. Severe acute respiratory syndrome. Nat Med. 2004;10(12):S88‐S97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Chan‐Yeung M, Xu RH. SARS: epidemiology. Respirology. 2003;8:S9‐S14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Lee J, Chowell G, Jung E. A dynamic compartmental model for the Middle East respiratory syndrome outbreak in the Republic of Korea: a retrospective analysis on control interventions and superspreading events. J Theor Biol. 2016;408:118‐126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Zaki AM, Van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med. 2012;367(19):1814‐1820. [DOI] [PubMed] [Google Scholar]
- 8. Lee JY, Kim Y‐J, Chung EH, et al. The clinical and virological features of the first imported case causing MERS‐CoV outbreak in South Korea, 2015. BMC Infect Dis. 2017;17(1):498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. of the International CSG. The species Severe acute respiratory syndrome‐related coronavirus: classifying 2019‐nCoV and naming it SARS‐CoV‐2. Nat Microbiol. 2020;5:536–544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Tan W, Zhao X, Ma X, Wang W, Niu P, Xu W. A novel coronavirus genome identified in a cluster of pneumonia cases—Wuhan, China 2019−2020. China CDC Weekly. 2020;2(4):61‐62. [PMC free article] [PubMed] [Google Scholar]
- 11. Lu R, Zhao X, Li J, et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet. 2020;395(10224):565‐574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Wong MC, Cregeen SJJ, Ajami NJ, Petrosino JF. Evidence of Recombination in Coronaviruses Implicating Pangolin Origins of NCoV‐2019 . 2020. 10.1101/2020.02.07.939207. [DOI] [Google Scholar]
- 13. Korber B, Fischer WM, Gnanakaran S, et al. Tracking changes in SARS‐CoV‐2 Spike: evidence that D614G increases infectivity of the COVID‐19 virus. Cell. 2020;182(4):812‐827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Vabret N, Britton GJ, Gruber C, et al. Immunology of COVID‐19: current state of the science. Immunity. 2020;52:910–941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Wu R, Wang L, Kuo H‐CD, et al. An update on current therapeutic drugs treating COVID‐19. Curr Pharmacol Rep. 2020;6:56–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Wang Y, Zhang D, Du G, et al. Remdesivir in adults with severe COVID‐19: a randomised, double‐blind, placebo‐controlled, multicentre trial. Lancet. 2020;395:1569–1578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Hillaker E, Belfer JJ, Bondici A, Murad H, Dumkow LE. Delayed initiation of remdesivir in a COVID‐19‐positive patient. Pharmacotherapy. 2020;40:592–598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Grein J, Ohmagari N, Shin D, et al. Compassionate use of remdesivir for patients with severe Covid‐19. N Engl J Med. 2020;382(24):2327‐2336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. RECOVERY Collaborative Group . Dexamethasone in hospitalized patients with Covid‐19 — preliminary report. N Engl J Med. 2020. 10.1056/nejmoa2021436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wu A, Peng Y, Huang B, et al. Genome composition and divergence of the novel coronavirus (2019‐nCoV) originating in China. Cell Host Microbe. 2020;27:325–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Zhou P, Yang X‐L, Wang X‐G, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270‐273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Cui J, Li F, Shi Z‐L. Origin and evolution of pathogenic coronaviruses. Nat Rev Microbiol. 2019;17(3):181‐192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Andersen KG, Rambaut A, Lipkin WI, Holmes EC, Garry RF. The proximal origin of SARS‐CoV‐2. Nat Med. 2020;26(4):450‐452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Wu F, Zhao S, Yu B, et al. A new coronavirus associated with human respiratory disease in China. Nature. 2020;579(7798):265‐269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Walls AC, Park Y‐J, Tortorici MA, Wall A, McGuire AT, Veesler D. Structure, function, and antigenicity of the SARS‐CoV‐2 spike glycoprotein. Cell. 2020;181:281–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Hoffmann M, Kleine‐Weber H, Schroeder S, et al. SARS‐CoV‐2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181:271–280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Jiang S, Hillyer C, Du L. Neutralizing antibodies against SARS‐CoV‐2 and other human coronaviruses. Trends Immunol. 2020;41:355–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Wu F, Wang A, Liu M, et al. Neutralizing antibody responses to SARS-CoV-2 in a COVID-19 recovered patient cohort and their implications. SSRN Electronic Journal. 10.2139/ssrn.3566211. [DOI] [Google Scholar]
- 29. Grifoni A, Weiskopf D, Ramirez SI, et al. Targets of T cell responses to SARS‐CoV‐2 coronavirus in humans with COVID‐19 disease and unexposed individuals. Cell. 2020;181:1489–1501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Braun J, Loyal L, Frentsch M, et al. Presence of Sars‐Cov‐2 Reactive T cells in Covid‐19 Patients and Healthy Donors . 2020. 10.1101/2020.04.17.20061440. [DOI] [PubMed] [Google Scholar]
- 31. Li F. Structure, function, and evolution of coronavirus spike proteins. Annu Rev Virol. 2016;3:237‐261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Wrapp D, Wang N, Corbett KS, et al. Cryo‐EM structure of the 2019‐nCoV spike in the prefusion conformation. Science. 2020;367(6483):1260‐1263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Pallesen J, Wang N, Corbett KS, et al. Immunogenicity and structures of a rationally designed prefusion MERS‐CoV spike antigen. Proc Natl Acad Sci. 2017;114(35):E7348‐E7357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Walls AC, Xiong X, Park Y‐J, et al. Unexpected receptor functional mimicry elucidates activation of coronavirus fusion. Cell. 2019;176(5):1026‐1039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Walls AC, Tortorici MA, Frenz B, et al. Glycan shield and epitope masking of a coronavirus spike protein observed by cryo‐electron microscopy. Nat Struct Mol Biol. 2016;23(10):899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Kirchdoerfer RN, Cottrell CA, Wang N, et al. Pre‐fusion structure of a human coronavirus spike protein. Nature. 2016;531(7592):118‐121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Tortorici MA, Veesler D. Structural insights into coronavirus entry. Adv virus Res. 2019;105:93‐116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Li Z, Tomlinson AC, Wong AH, et al. The human coronavirus HCoV‐229E S‐protein structure and receptor binding. Elife. 2019;8:e51230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Wong AH, Tomlinson AC, Zhou D, et al. Receptor‐binding loops in alphacoronavirus adaptation and evolution. Nat Commun. 2017;8(1):1‐10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Wu K, Li W, Peng G, Li F. Crystal structure of NL63 respiratory coronavirus receptor‐binding domain complexed with its human receptor. Proc Natl Acad Sci. 2009;106(47):19970‐19974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Watanabe Y, Allen JD, Wrapp D, McLellan JS, Crispin M. Site‐Specific Analysis of the SARS‐CoV‐2 Glycan Shield . 2020. 10.1101/2020.03.26.010322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Guo L, Ren L, Yang S, et al. Profiling early humoral response to diagnose novel coronavirus disease (COVID‐19). Clin Infect Dis. 2020;71:778–785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Theel ES, Slev P, Wheeler S, Couturier MR, Wong SJ, Kadkhoda K. The role of antibody testing for SARS‐CoV‐2: is there one? J Clin Microbiol. 2020;58:e00797‐20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Guo C, Li B, Ma H, et al. Single‐cell analysis of two severe COVID‐19 patients reveals a monocyte‐associated and tocilizumab‐responding cytokine storm. Nat Commun. 2020;11(1). 10.1038/s41467-020-17834-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Wen W, Su W, Tang H, et al. Immune cell profiling of COVID‐19 patients in the recovery stage by single‐cell sequencing. Cell Discov. 2020;6(1):1‐18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Ju B, Zhang Q, Ge J, et al. Human neutralizing antibodies elicited by SARS‐CoV‐2 infection. Nature. 2020:1‐8. [DOI] [PubMed] [Google Scholar]
- 47. Bao L, Deng W, Gao H, et al. Reinfection Could Not Occur in SARS‐CoV‐2 Infected Rhesus Macaques . 2020. 10.1101/2020.03.13.990226. [DOI] [Google Scholar]
- 48. Patel R, Babady E, Theel ES, et al. Report from the American Society for Microbiology COVID‐19 international summit, 23 march 2020: value of diagnostic testing for SARS–CoV‐2/COVID‐19. Am Soc Microbiol. 2020;11:e00722‐20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Lu B, Huang Y, Huang L, et al. Effect of mucosal and systemic immunization with virus‐like particles of severe acute respiratory syndrome coronavirus in mice. Immunology. 2010;130(2):254‐261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Shim B‐S, Stadler K, Nguyen HH, et al. Sublingual immunization with recombinant adenovirus encoding SARS‐CoV spike protein induces systemic and mucosal immunity without redirection of the virus to the brain. Virology J. 2012;9(1):1‐9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. To KK‐W, Tsang OT‐Y, Leung W‐S, et al. Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS‐CoV‐2: an observational cohort study. Lancet Infect Dis. 2020;20:565–574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Tian X, Li C, Huang A, et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus‐specific human monoclonal antibody. Emerg Microbes Infect. 2020;9(1):382‐385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Tai W, He L, Zhang X, et al. Characterization of the receptor‐binding domain (RBD) of 2019 novel coronavirus: implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell Mol Immunol. 2020;17:613‐620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Chen W‐H, Hotez PJ, Bottazzi ME. Potential for developing a SARS‐CoV receptor‐binding domain (RBD) recombinant protein as a heterologous human vaccine against coronavirus infectious disease (COVID)‐19. Hum Vaccines Immunother. 2020;16:1239‐1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Zhou Y, Yang Y, Huang J, Jiang S, Du L. Advances in MERS‐CoV vaccines and therapeutics based on the receptor‐binding domain. Viruses. 2019;11(1):60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Zhu Z, Chakraborti S, He Y, et al. Potent cross‐reactive neutralization of SARS coronavirus isolates by human monoclonal antibodies. Proc Natl Acad Sci. 2007;104(29):12123‐12128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Rockx B, Corti D, Donaldson E, et al. Structural basis for potent cross‐neutralizing human monoclonal antibody protection against lethal human and zoonotic severe acute respiratory syndrome coronavirus challenge. J virology. 2008;82(7):3220‐3235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Sui J, Li W, Murakami A, et al. Potent neutralization of severe acute respiratory syndrome (SARS) coronavirus by a human mAb to S1 protein that blocks receptor association. Proc Natl Acad Sci. 2004;101(8):2536‐2541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. He Y, Li J, Li W, Lustigman S, Farzan M, Jiang S. Cross‐neutralization of human and palm civet severe acute respiratory syndrome coronaviruses by antibodies targeting the receptor‐binding domain of spike protein. J Immunol. 2006;176(10):6085‐6092. [DOI] [PubMed] [Google Scholar]
- 60. Pinto D, Park Y‐J, Beltramello M, et al. Structural and Functional Analysis of a Potent Sarbecovirus Neutralizing Antibody . 2020. 10.1101/2020.04.07.023903. [DOI] [Google Scholar]
- 61. Yuan M, Wu NC, Zhu X, et al. A highly conserved cryptic epitope in the receptor binding domains of SARS‐CoV‐2 and SARS‐CoV. Science. 2020;368(6491):630‐633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Chen X, Li R, Pan Z, et al. Human monoclonal antibodies block the binding of SARS‐CoV‐2 spike protein to angiotensin converting enzyme 2 receptor. Cell Mol Immunol. 2020;17:647‐649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Wölfel R, Corman VM, Guggemos W, et al. Virological assessment of hospitalized patients with COVID‐2019. Nature. 2020;581(7809):465‐469. [DOI] [PubMed] [Google Scholar]
- 64. Wang C, Li W, Drabek D, et al. A human monoclonal antibody blocking SARS‐CoV‐2 infection. Nat Commun. 2020;11(1):1‐6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Wu Y, Li C, Xia S, et al. Identification of human single‐domain antibodies against SARS‐CoV‐2. Cell host microbe. 2020;27:891–898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Wrapp D, De Vlieger D, Corbett KS, et al. Structural basis for potent neutralization of betacoronaviruses by single‐domain camelid antibodies. Cell. 2020;181:1004–1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Lei C, Qian K, Li T, et al. Neutralization of SARS‐CoV‐2 spike pseudotyped virus by recombinant ACE2‐Ig. Nat Commun. 2020;11(1):1‐5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Li Y, Wang H, Tang X, et al. SARS-CoV-2 and three related coronaviruses utilize multiple ACE2 orthologs and are potently blocked by an improved ACE2-Ig. J Virol. 2020. 10.1128/jvi.01283-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Okba NM, Müller MA, Li W, et al. Severe acute respiratory syndrome coronavirus 2‐specific antibody responses in coronavirus disease 2019 patients. Emerg Infect Dis. 2020;26(7):1478–1488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Zhou F, Yu T, Du R, et al. Clinical course and risk factors for mortality of adult inpatients with COVID‐19 in Wuhan, China: a retrospective cohort study. Lancet. 2020;395:1054–1062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Li K, Chen D, Chen S, et al. Radiographic Findings and Other Predictors in Adults with Covid‐19. MedRxiv; 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Zhang L, Zhang F, Yu W, et al. Antibody responses against SARS coronavirus are correlated with disease outcome of infected individuals. J Med Virol. 2006;78(1):1‐8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Liu L, Wei Q, Lin Q, et al. Anti‐spike IgG causes severe acute lung injury by skewing macrophage responses during acute SARS-CoV infection. JCI Insight. 2019;4(4). 10.1172/jci.insight.123158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Wan Y, Shang J, Sun S, et al. Molecular mechanism for antibody-dependent enhancement of coronavirus entry. J Virol. 2019;94(5). 10.1128/jvi.02015-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Lambert P‐H, Ambrosino DM, Andersen SR, et al. Consensus summary report for CEPI/BC March 12‐13, 2020 meeting: assessment of risk of disease enhancement with Covid‐19 vaccines. Vaccine. 2020;38:4783–4791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Padron‐Regalado E. Vaccines for SARS‐CoV‐2: lessons from other coronavirus strains. Infect Dis Ther. 2020;9:255‐274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Choi AH, Basu M, McNeal MM, Clements JD, Ward RL. Antibody‐independent protection against rotavirus infection of mice stimulated by intranasal immunization with chimeric VP4 or VP6 protein. J Virol. 1999;73(9):7574‐7581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Diao B, Wang C, Tan Y, et al. Reduction and functional exhaustion of T cells in patients with coronavirus disease 2019 (COVID‐19). Front Immunol. 2020;11:827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Wan S, Yi Q, Fan S, et al. Characteristics of Lymphocyte Subsets and Cytokines in Peripheral Blood of 123 Hospitalized Patients with 2019 Novel Coronavirus Pneumonia (NCP) . 2020. 10.1101/2020.02.10.20021832. [DOI] [Google Scholar]
- 80. Sun D‐w, Zhang D, Tian R‐h, et al. The underlying changes and predicting role of peripheral blood inflammatory cells in severe COVID‐19 patients: a sentinel? Clin Chim Acta. 2020;508:122–129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Huang C, Wang Y, Li X, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. lancet. 2020;395(10223):497‐506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Liu Y, Zhang C, Huang F, et al. Elevated plasma level of selective cytokines in COVID‐19 patients reflect viral load and lung injury. Nat Sci Rev. 2020;7:1003–1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Buonaguro FM, Ascierto PA, Morse GD, et al. Covid‐19: time for a paradigm change. Rev Med Virol. 2020;30:e2134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Li CK‐f, Wu H, Yan H, et al. T cell responses to whole SARS coronavirus in humans. J Immunol. 2008;181(8):5490‐5500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Channappanavar R, Fett C, Zhao J, Meyerholz DK, Perlman S. Virus‐specific memory CD8 T cells provide substantial protection from lethal severe acute respiratory syndrome coronavirus infection. J Virol. 2014;88(19):11034‐11044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Tang F, Quan Y, Xin Z‐T, et al. Lack of peripheral memory B cell responses in recovered patients with severe acute respiratory syndrome: a six‐year follow‐up study. J Immunol. 2011;186(12):7264‐7268. [DOI] [PubMed] [Google Scholar]
- 87. Peng H, Yang L‐t, Wang L‐y, et al. Long‐lived memory T lymphocyte responses against SARS coronavirus nucleocapsid protein in SARS‐recovered patients. Virology. 2006;351(2):466‐475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Fan Y‐Y, Huang Z‐T, Li L, et al. Characterization of SARS‐CoV‐specific memory T cells from recovered individuals 4 years after infection. Arch Virol. 2009;154(7):1093‐1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Ahmed SF, Quadeer AA, McKay MR. Preliminary identification of potential vaccine targets for the COVID‐19 coronavirus (SARS‐CoV‐2) based on SARS‐CoV immunological studies. Viruses. 2020;12(3):254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Vashi Y, Jagrit V, Kumar S. Understanding the B and T cells Epitopes of Spike Protein of Severe Respiratory Syndrome Coronavirus‐2: a Computational Way to Predict the Immunogens . 2020. 10.1101/2020.04.08.013516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Li J, Wang J, Kang AS, Sacitharan PK. Mapping the T cell response to COVID‐19. Signal Transduct Target Ther. 2020;5(1):112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Ni L, Ye F, Cheng M‐L, et al. Detection of SARS‐CoV‐2‐specific humoral and cellular immunity in COVID‐19 convalescent individuals. Immunity. 2020;52:971–977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Weiskopf D, Schmitz KS, Raadsen MP, et al. Phenotype and kinetics of SARS‐CoV‐2‐specific T cells in COVID‐19 patients with acute respiratory distress syndrome. Sci Immunol. 2020;5(48):eabd2071. 10.1126/sciimmunol.abd2071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Sekine T, Perez‐Potti A, Rivera‐Ballesteros O, et al. Robust T cell immunity in convalescent individuals with asymptomatic or mild COVID-19. Cell. 2020;183(1):158–168.e14. 10.1016/j.cell.2020.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Tseng C‐T, Sbrana E, Iwata‐Yoshikawa N, et al. Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PLoS One. 2012;7(4):e35421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Bolles M, Deming D, Long K, et al. A double‐inactivated severe acute respiratory syndrome coronavirus vaccine provides incomplete protection in mice and induces increased eosinophilic proinflammatory pulmonary response upon challenge. J Virol. 2011;85(23):12201‐12215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Agrawal AS, Tao X, Algaissi A, et al. Immunization with inactivated Middle East Respiratory Syndrome coronavirus vaccine leads to lung immunopathology on challenge with live virus. Hum Vaccin Immunother. 2016;12(9):2351‐2356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Zhao J, Li K, Wohlford‐Lenane C, et al. Rapid generation of a mouse model for Middle East respiratory syndrome. Proc Natl Acad Sci. 2014;111(13):4970‐4975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Houser KV, Broadbent AJ, Gretebeck L, et al. Enhanced inflammation in New Zealand white rabbits when MERS‐CoV reinfection occurs in the absence of neutralizing antibody. PLoS Pathog. 2017;13(8):e1006565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Sun J, He W‐T, Wang L, et al. COVID‐19: epidemiology, evolution, and cross‐disciplinary perspectives. Trends Mol Med. 2020;26:483–495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Ye T, Zhong Z, García‐Sastre A, Schotsaert M, De Geest BG. Current status of COVID‐19 (pre)clinical vaccine development. Angew Chem Int Ed. 2020. 10.1002/anie.202008319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Le TT, Andreadakis Z, Kumar A, et al. The COVID‐19 vaccine development landscape. Nat Rev Drug Discov. 2020;19(5):305‐306. [DOI] [PubMed] [Google Scholar]
- 103. Zhao P, Ke J‐S, Qin Z‐L, et al. DNA vaccine of SARS‐Cov S gene induces antibody response in mice. Acta Biochim Biophys Sin. 2004;36(1):37‐41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Muthumani K, Falzarano D, Reuschel EL, et al. A synthetic consensus anti–spike protein DNA vaccine induces protective immunity against Middle East respiratory syndrome coronavirus in nonhuman primates. Sci Transl Med. 2015;7(301):301ra132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Bisht H, Roberts A, Vogel L, et al. Severe acute respiratory syndrome coronavirus spike protein expressed by attenuated vaccinia virus protectively immunizes mice. Proc Natl Acad Sci. 2004;101(17):6641‐6646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Chen Z, Zhang L, Qin C, et al. Recombinant modified vaccinia virus Ankara expressing the spike glycoprotein of severe acute respiratory syndrome coronavirus induces protective neutralizing antibodies primarily targeting the receptor binding region. J Virol. 2005;79(5):2678‐2688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Bukreyev A, Lamirande EW, Buchholz UJ, et al. Mucosal immunisation of African green monkeys (Cercopithecus aethiops) with an attenuated parainfluenza virus expressing the SARS coronavirus spike protein for the prevention of SARS. Lancet. 2004;363(9427):2122‐2127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Kam YW, Kien F, Roberts A, et al. Antibodies against trimeric S glycoprotein protect hamsters against SARS‐CoV challenge despite their capacity to mediate FcγRII‐dependent entry into B cells in vitro. Vaccine. 2007;25(4):729‐740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Vennema H, De Groot R, Harbour D, et al. Early death after feline infectious peritonitis virus challenge due to recombinant vaccinia virus immunization. J Virol. 1990;64(3):1407‐1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. Czub M, Weingartl H, Czub S, He R, Cao J. Evaluation of modified vaccinia virus Ankara based recombinant SARS vaccine in ferrets. Vaccine. 2005;23(17‐18):2273‐2279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Weingartl H, Czub M, Czub S, et al. Immunization with modified vaccinia virus Ankara‐based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets. J Virol. 2004;78(22):12672‐12676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Honda‐Okubo Y, Barnard D, Ong CH, Peng B‐H, Tseng C‐TK, Petrovsky N. Severe acute respiratory syndrome‐associated coronavirus vaccines formulated with delta inulin adjuvants provide enhanced protection while ameliorating lung eosinophilic immunopathology. J Virol. 2015;89(6):2995‐3007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Yang Z‐y, Kong W‐p, Huang Y, et al. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature. 2004;428(6982):561‐564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. He Y, Li J, Heck S, Lustigman S, Jiang S. Antigenic and immunogenic characterization of recombinant baculovirus‐expressed severe acute respiratory syndrome coronavirus spike protein: implication for vaccine design. J Virol. 2006;80(12):5757‐5767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Coleman CM, Liu YV, Mu H, et al. Purified coronavirus spike protein nanoparticles induce coronavirus neutralizing antibodies in mice. Vaccine. 2014;32(26):3169‐3174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Draft landscape of COVID‐19 candidate vaccines [updated 2020]. https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines.
- 117. Folegatti PM, Ewer KJ, Aley PK, et al. Safety and immunogenicity of the ChAdOx1 nCoV‐19 vaccine against SARS‐CoV‐2: a preliminary report of a phase 1/2, single‐blind, randomised controlled trial. Lancet. 2020;396:467–478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. van Doremalen N, Lambe T, Spencer A, et al. ChAdOx1 NCoV‐19 Vaccination Prevents SARS‐CoV‐2 Pneumonia in Rhesus Macaques . 2020. 10.1101/2020.05.13.093195. [DOI] [PubMed] [Google Scholar]
- 119. Zhu F‐C, Li Y‐H, Guan X‐H, et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type‐5 vectored COVID‐19 vaccine: a dose‐escalation, open‐label, non‐randomised, first‐in‐human trial. Lancet. 2020;395:1845–1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Corbett KS, Edwards DK, Leist SR, et al. SARS‐CoV‐2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature. 2020. 10.1038/s41586-020-2622-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Walsh EE, Frenck R, Falsey AR, et al. RNA‐based COVID‐19 Vaccine BNT162b2 Selected for a Pivotal Efficacy Study. 2020. 10.1101/2020.08.17.20176651. [DOI] [Google Scholar]
- 122. Keech C, Albert G, Cho I, et al. Phase 1–2 trial of a SARS‐CoV‐2 recombinant spike protein nanoparticle vaccine. N Engl Med. 2020. 10.1056/nejmoa2026920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Jackson LA., Anderson EJ, Rouphael NG, et al. An mRNA vaccine against SARS‐CoV‐2—preliminary report. N Engl J Med. 2020. 10.1056/nejmoa2022483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Vogel A, Kanevsky I, Che Y, et al. A Prefusion SARS‐CoV‐2 Spike RNA Vaccine is Highly Immunogenic and Prevents Lung Infection in Non‐Human Primates . 2020. 10.1101/2020.09.08.280818. [DOI] [Google Scholar]
- 125. Tian J‐H, Patel N, Haupt R, et al. SARS‐CoV‐2 Spike Glycoprotein Vaccine Candidate NVX‐CoV2373 Elicits Immunogenicity in Baboons and Protection in Mice . 2020. 10.1101/2020.06.29.178509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Guebre‐Xabier M, Patel N, Tian J‐H, et al. NVX‐CoV2373 Vaccine Protects Cynomolgus Macaque Upper and Lower Airways Against SARS‐CoV‐2 Challenge . 2020; 10.1101/2020.08.18.256578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Du L, He Y, Zhou Y, Liu S, Zheng B‐J, Jiang S. The spike protein of SARS‐CoV—a target for vaccine and therapeutic development. Nat Rev Microbiol. 2009;7(3):226‐236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Zhu X, Liu Q, Du L, Lu L, Jiang S. Receptor‐binding domain as a target for developing SARS vaccines. J Thorac Dis. 2013;5(suppl 2):S142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Lan J, Yao Y, Deng Y, et al. Recombinant receptor binding domain protein induces partial protective immunity in rhesus macaques against Middle East respiratory syndrome coronavirus challenge. EBioMedicine. 2015;2(10):1438‐1446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Jiang S, He Y, Liu S. SARS vaccine development. Emerg Infect Dis. 2005;11(7):1016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Yang Y, Deng Y, Wen B, et al. The amino acids 736–761 of the MERS‐CoV spike protein induce neutralizing antibodies: implications for the development of vaccines and antiviral agents. Viral Immunol. 2014;27(10):543‐550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Tai W, Wang Y, Fett CA, et al. Recombinant receptor‐binding domains of multiple Middle East respiratory syndrome coronaviruses (MERS‐CoVs) induce cross‐neutralizing antibodies against divergent human and camel MERS‐CoVs and antibody escape mutants. J Virol. 2017;91(1):e01651‐16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. He Y, Zhou Y, Siddiqui P, Jiang S. Inactivated SARS‐CoV vaccine elicits high titers of spike protein‐specific antibodies that block receptor binding and virus entry. Biochem Biophys Res Commun. 2004;325(2):445‐452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. He Y, Zhu Q, Liu S, et al. Identification of a critical neutralization determinant of severe acute respiratory syndrome (SARS)‐associated coronavirus: importance for designing SARS vaccines. Virology. 2005;334(1):74‐82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. He Y, Zhou Y, Liu S, et al. Receptor‐binding domain of SARS‐CoV spike protein induces highly potent neutralizing antibodies: implication for developing subunit vaccine. Biochem Biophys Res Commun. 2004;324(2):773‐781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Du L, Zhao G, He Y, et al. Receptor‐binding domain of SARS‐CoV spike protein induces long‐term protective immunity in an animal model. Vaccine. 2007;25(15):2832‐2838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. Nyon MP, Du L, Tseng C‐TK, et al. Engineering a stable CHO cell line for the expression of a MERS‐coronavirus vaccine antigen. Vaccine. 2018;36(14):1853‐1862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Du L, Zhao G, Lin Y, et al. Priming with rAAV encoding RBD of SARS‐CoV S protein and boosting with RBD‐specific peptides for T cell epitopes elevated humoral and cellular immune responses against SARS‐CoV infection. Vaccine. 2008;26(13):1644‐1651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Du L, He Y, Wang Y, et al. Recombinant adeno‐associated virus expressing the receptor‐binding domain of severe acute respiratory syndrome coronavirus S protein elicits neutralizing antibodies: implication for developing SARS vaccines. Virology. 2006;353(1):6‐16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Du L, Zhao G, Lin Y, et al. Intranasal vaccination of recombinant adeno‐associated virus encoding receptor‐binding domain of severe acute respiratory syndrome coronavirus (SARS‐CoV) spike protein induces strong mucosal immune responses and provides long‐term protection against SARS‐CoV infection. J Immunol. 2008;180(2):948‐956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Zhi Y, Kobinger GP, Jordan H, et al. Identification of murine CD8 T cell epitopes in codon‐optimized SARS‐associated coronavirus spike protein. Virology. 2005;335(1):34‐45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Sahin U, Muik A, Derhovanessian E, et al. Concurrent Human Antibody and TH1 type T‐cell Responses Elicited by a COVID‐19 RNA Vaccine . 2020; 10.1101/2020.07.17.20140533. [DOI] [Google Scholar]
- 143. Mulligan MJ, Lyke KE, Kitchin N, et al. Phase 1/2 Study to Describe the Safety and Immunogenicity of a COVID‐19 RNA Vaccine Candidate (BNT162b1) in Adults 18 to 55 Years of Age: Interim Report. 2020. [Google Scholar]
- 144. Cheng Y, He B, Yang J, et al. Crystal structure of the S1 subunit N‐terminal domain from DcCoV UAE‐HKU23 spike protein. Virology. 2019;535:74‐82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Qian Z, Ou X, Góes LGB, et al. Identification of the receptor‐binding domain of the spike glycoprotein of human betacoronavirus HKU1. J Virol. 2015;89(17):8816‐8827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Jiaming L, Yanfeng Y, Yao D, et al. The recombinant N‐terminal domain of spike proteins is a potential vaccine against Middle East respiratory syndrome coronavirus (MERS‐CoV) infection. Vaccine. 2017;35(1):10‐18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Chen Y, Lu S, Jia H, et al. A novel neutralizing monoclonal antibody targeting the N‐terminal domain of the MERS‐CoV spike protein. Emerg microbes Infect. 2017;6(1):1‐7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148. Adney DR, Wang L, van Doremalen N, et al. Efficacy of an adjuvanted Middle East respiratory syndrome coronavirus spike protein vaccine in dromedary camels and alpacas. Viruses. 2019;11(3):212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Liu R‐Y, Wu L‐Z, Huang B‐J, et al. Adenoviral expression of a truncated S1 subunit of SARS‐CoV spike protein results in specific humoral immune responses against SARS‐CoV in rats. Virus Res. 2005;112(1‐2):24‐31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Wang Y, Tai W, Yang J, et al. Receptor‐binding domain of MERS‐CoV with optimal immunogen dosage and immunization interval protects human transgenic mice from MERS‐CoV infection. Hum Vaccin Immunother. 2017;13(7):1615‐1624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Kim E, Erdos G, Huang S, et al. Microneedle array delivered recombinant coronavirus vaccines: immunogenicity and rapid translational development. EBioMedicine. 2020;55:102743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Elshabrawy HA, Coughlin MM, Baker SC, Prabhakar BS. Human monoclonal antibodies against highly conserved HR1 and HR2 domains of the SARS‐CoV spike protein are more broadly neutralizing. PLoS One. 2012;7(11):e50366. 10.1371/journal.pone.0050366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Zhou Y, Jiang S, Du L. Prospects for a MERS‐CoV spike vaccine. Expet Rev Vaccine. 2018;17(8):677‐686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Shang W, Yang Y, Rao Y, Rao X. The outbreak of SARS‐CoV‐2 pneumonia calls for viral vaccines. NPJ Vaccines. 2020;5(1):1‐3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155. Chan JF‐W, Kok K‐H, Zhu Z, et al. Genomic characterization of the 2019 novel human‐pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg Microbes Infect. 2020;9(1):221‐236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Wang N, Shang J, Jiang S, Du L. Subunit vaccines against emerging pathogenic human coronaviruses. Front Microbiol. 2020;11:298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157. Lu L, Liu Q, Zhu Y, et al. Structure‐based discovery of Middle East respiratory syndrome coronavirus fusion inhibitor. Nat Commun. 2014;5(1):1‐12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Guo Y, Sun S, Wang K, Zhang S, Zhu W, Chen Z. Elicitation of immunity in mice after immunization with the S2 subunit of the severe acute respiratory syndrome coronavirus. DNA Cell Biol. 2005;24(8):510‐515. [DOI] [PubMed] [Google Scholar]
- 159. Zhang J, Zeng H, Gu J, Li H, Zheng L, Zou Q. Progress and prospects on vaccine development against SARS‐CoV‐2. Vaccines. 2020;8(2):153. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data openly available in a public repository that issues datasets with DOIs.