
Hipótese da origem única

Hipótese da origem única (no inglês recent single-origin hypothesis, ou RSOH), também chamada de Modelo "Fora da África" (no Inglês "Out of Africa", ou OOA), Hipótese da Substituição ou Origem Recente Africana (no inglês Recent African Origin, ou RAO) é o modelo da Paleoantropologia mais amplamente aceito para a origem humana, corroborado por evidências fósseis, genéticas e linguísticas, que indicam que todos os seres humanos hoje vivos descendem de um grupo de Homo sapiens originado na África, sendo as estimativas de datas de coalescência mais atualizadas, obtidas a partir de pesquisas genéticas, de há 143 mil a 190 mil anos para as linhagens materna e paterna, respectivamente,[1][2] porém com que é chamado Humano Anatomicamente Moderno (no inglês Anatomically Modern Human, ou AMH) tendo se originado há entre 100 e 300 mil anos (vide Tabela 1).[3] Os descendentes desse grupo posteriormente se dispersaram pelo mundo em diferentes ondas migratórias, constituindo as linhagens que deram origem aos povos viventes de todos os continentes, tendo a maior e mais significativa dessas ondas migratórias se iniciado por volta de há 60 a 75 milhares de anos.[3][4][5]
O modelo original propunha que esses Homo sapiens modernos migrantes substituiram outras espécies do gênero Homo já presentes fora do continente africano, como Homo neanderthalensis e Homo denisova, porém, hoje se tem conhecimento que ocorreu um certo grau hibridações entre Homo sapiens e as outros hominínios "não-modernos" dentro e fora da África, que faz com que os humanos viventes tenham uma pequena porcentagem de material genético advindo dessas outras espécies.[6][7]
Atualmente, a hipótese mais aceita é que o homem moderno saiu da África e colonizou outras partes do mundo, em várias ondas migratórias, corroborada por evidências fósseis, moleculares, microbiológicas. Novas análises de sequências do cromossomo Y demonstraram uma grande diversidade entre as linhas paternas no sul e sudeste da Ásia. Esta evidência corrobora outro estudo feito anteriormente, em que, por meio de análises feitas usando um dente de homem moderno, o sudeste da Ásia foi identificado como um dos primeiros centros de dispersão humana. Análises feitas no DNA mitocondrial apontaram o sudoeste europeu como outro centro de dispersão secundária do homem moderno há 40.000 anos. Dados moleculares e linguísticos indicam Beringia como outro importante centro secundário de dispersão.[3]
"Em um sentido genético, todos nesse planeta se parecem africanos."Original (em inglês): "In a genetic sense, everyone on this planet looks like an African."— Svante Pääbo,Hoffecker, J. F. (2017). Modern humans: Their African origin and global dispersal. Columbia University Press. (em inglês)
Humanos Modernos
[editar | editar código-fonte]Os Homo sapiens modernos compartilham algumas características esqueléticas que podem ser identificadas no registro fóssil, como neurocrânio alto e arredondado, uma face pequena acomodada abaixo dele e uma arcada supraciliar pequena e dividida em duas partes;[8] essas delimitam o que é chamado Humano Anatomicamente Moderno (no inglês Anatomically Modern Human, ou AMH), que inclui todos os seres humanos hoje viventes.[7] Esses traços surgiram e se estabeleceram há entre 300 e 150 milhares de anos na África (vide Tabela 1)[3] e foram exportados para o resto do mundo por expansões migratórias de humanos que os continham, acontecendo com maior significância a partir de há aproximadamente 60 mil anos.[9] Em contraste, pode se chamar Homo sapiens 'arcaico', ou humanos arcaicos, aqueles fósseis de hominídeos que se encaixariam na linhagem humana direta entre a divergência a partir de Homo heidelbergensis (nosso provável ancestral comum com H. neanderthalensis e H. denisova) há aproximadamente 500 mil anos[3] e o estabelecimento dessas características diagnósticas de um humano moderno, porém é recorrente o uso desse termo para se referir a outras espécies do gênero Homo.[7]
Além da caracterização anatômica dos seres humanos modernos, há hipóteses que propõem que a evolução de Homo sapiens se deu em duas etapas separadas, primeiramente com o estabelecimento gradual dessas características morfológicas que são vistas no registro fóssil e, posteriormente, após provavelmente 100 mil anos antes do presente, o desenvolvimento do comportamento moderno e faculdades cognitivas observadas nos humanos viventes. Essa hipótese se apoia principalmente em evidências arqueológicas que indicam um surgimento tardio de comportamentos simbólicos, em relação ao surgimento da morfologia moderna.[10] Richard G. Klein integrou os dados arqueológicos e fósseis e usou os dados para argumentar essa emergência do comportamento moderno completo pode ser o que possibilitou a expansão humana por todo o mundo, sendo essa chamada a "hipótese neural" da paleoantropologia.[11][3] Também há argumentos apontando que as evidências fósseis e arqueológicas indicam uma evolução gradual tanto da morfologia, quanto da cognição humanas, essas evidências foram sistematizadas em um artigo de alto impacto publicado por McBrearty & Brooks em 2000.[12]
Morfologia (Fósseis)
[editar | editar código-fonte]
As descrições operacionais usada para definir humanos modernos osteologicamente visam abranger o máximo de variação possível, porém elas sempre serão falhas e poderão não englobar exceções, portanto é necessário ressaltar que todos os seres humanos viventes estão inclusos na categoria de humanos anatomicamente modernos, mesmo suas morfologias não estejam dentro dos parâmetros diagnósticos usados na paleoantropologia.[7][13]
Em linhas gerais o esqueleto de um Humano Anatomicamente Moderno pode ser descrito formalmente por algumas medidas métricas da morfologia do Crânio e Pós-crânio,[14][15][13][16] propostas inicialmente por Michael Day e Christopher Stringer no Congresso Internacional de Paleontologia Humana de 1982, sediado em Nice.[17]
Além disso, é possível se distinguir os fósseis de Homo sapiens dos de outros hominínios a partir do padrão de crescimento dos dentes, o qual está relacionado ao padrão de desenvolvimento ontogenético retardado em H. sapiens, que, por sua vez, está relacionado ao desenvolvimento das faculdades cognitivas características dessa espécie.[18] Porém, esse padrão já é observado mesmo em fósseis que ainda apresentam um mosaico de características modernas e arcaicas, como os de 300 mil anos de Djebel Irhoud, no Marrocos.[19]
Comportamento (Artefatos)
[editar | editar código-fonte]Como comportamentos não se fossilizam, as melhores evidências que temos de sua evolução são os artefatos gerados por eles, no caso de um comportamento humano considerado moderno e associado à uma capacidade cognitiva equivalente à dos seres humanos viventes, podendo esses artefatos estarem associados a características como exibição social, trocas de longa distância, notações digitais (engraves) ou capacidades artísticas.[10][11][12]
Achados arqueológicos da Caverna Blombos datados em aproximadamente 77 mil anos antes do presente, na África do Sul, se destacam entre os artefatos que podem indicar um comportamento complexo, entre eles estão conchas perfuradas, provavelmente usadas para confecção de colares e outras ornamentações corporais, e um pedaço de ocre vermelho com entalhes geométricos.[20][21] Pontas ósseas farpadas encontradas em Catanda, Congo, e datadas em 90 mil anos também são uma evidência do possível surgimento de um comportamento moderno.[22]
-
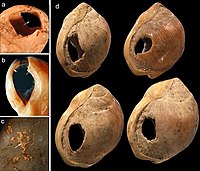 Conchas perfuradas, Caverna Blombos, África do Sul.
Conchas perfuradas, Caverna Blombos, África do Sul. -
 Exemplo de pontas ósseas farpadas
Exemplo de pontas ósseas farpadas -
 Gravações em fragmentos do ovos de avestruz, Diepkloof, África do Sul
Gravações em fragmentos do ovos de avestruz, Diepkloof, África do Sul

._Oban,_Argyll%27s_P.S.A.S._Wellcome_M0015020.jpg)

Histórico da Hipótese
[editar | editar código-fonte]Até a década de 50 a paleoantropologia enfrentava o problema da ausência de métodos precisos e confiáveis para se datar os restos fósseis e até a década de 60 os registros fósseis de H. sapiens mais antigos pertenciam a sítios na Europa ou na Ásia. Além disso, a paleoantropologia possuía fortes vieses raciais, com uma visão de atraso da África em relação à Europa.[23][3]
Em 1967, foram descobertos, por Richard Leakey e sua equipe, restos fósseis humanos no que hoje é chamado de sítio Omo-Kibish, no Rio Omo, Etiópia.[3] O crânio fóssil conhecido como Omo-Kibish I foi datado por Karl Butzer em cerca de 130 mil anos, até então mais antigo que qualquer evidência prévia de humanos presentes em qualquer outra região do mundo.[24] Já durante os anos 70 e 80 mais restos humanos foram encontrados e/ou datados, no leste e sul da África, em faixas temporais que antecedem a presença de seres humanos modernos no resto do mundo, incluindo os de Border Cave (África do Sul), Florisbad (África do Sul), Eliye Springs (Quénia) e Cavernas Rio Klassies (África do Sul), vide Tabela 1.[25]
A hipótese da Origem Recente Africana só tomou corpo em meados dos anos 80, principalmente pelo trabalho dos paleoantropólogos Günter Bräuer, na Alemanha, e Chris Stringer e Michael Day, no Reino Unido; os quais, no Congresso Internacional de Paleontologia Humana de 1982, oficialmente apresentaram o modelo e argumentos para que os fósseis encontrados no sul e no leste africano poderiam caracterizar membros de Homo sapiens, na qual tembém proporam uma maneira sistemática de diagnosticar um humano moderno através de traços anatômicos esqueléticos.[17][10][3] A noção de uma origem única na África surgiu como um contraste principalmente à hipótese de Evolução Multirregional, que constituía o modelo dominante para explicar o surgimento humano por todo o planeta.[10]
A partir de 1987 o debate das origens humanas entrou em uma nova instância, com a publicação de um trabalho por Rebecca Cann, Mark Stoneking e Alan Wilson, o qual analisou sequências de DNA mitocondrial (mtDNA) de 147 mulheres ao redor do mundo, representando diferentes linhagens. O trabalho apontou que todas a linhagens de fêmeas humana hoje viventes derivaram de uma ancestral comum há aproximadamente 200 mil anos.[26] A partir desse estudo se popularizaram termos como "Eva Mitocondrial" ou "Eva Africana" para se referir a essa ancestral comum de todas as mulheres humana viventes.[3]

Desde a publicação dessa primeira análise de mtDNA as análises genéticas passaram a dominar as pesquisas na área, em sua maior parte corroborando o modelo ou fazendo adições a ele, com a realização subsequente de análises de cromossomo Y, DNA ancestral (aDNA),[28] e mais recentemente análises genômicas de populações e de material genético de microorganismos comensais ou patogênicos que sofreram coevolução com os humanos.[29][30][31][32][33]
Até 2010, a discussão sobre hibridação entre humanos modernos e Neandertais se baseava em traços anatômicos observados no registro fóssil e carecia de bases sólidas, com o trabalho publicado em 2003 por Erik Trinkaus e pares, que aprontava características Neandertais em um fóssil humano encontrado em Oase, Romênia.[34] Porém, em 2010 foram publicados dois trabalhos independentes que, através de análises genéticas genômicas possibilitadas pelo método de "Sequenciamento de Nova Geração", apontavam a presença de cerca de 2,5% de DNA Neandertal nas populações Humana não Africanas e aproximadamente 5% de DNA Denisovano em populações nativas da Austrália e Nova Guiné.[35][36] Essas publicações abriram uma nova epoca no debate sobre as origens humanas, tirando a dominânancia da hipótese de uma origem exclusivamente africana e deslocando o consenso para o que pode se considerar o modelo "Origem Recente Africana + Hibridação".[37]
Evidências
[editar | editar código-fonte]A Origem Recente Africana, ou RAO, do inglês Recent African Origin, e, posteriormente, a dispersão global dos humanos modernos tem três pilares de evidências: a) dados fósseis de humanos modernos e não-modernos, b) análise genética, estudo do genoma de pessoas viventes atualmente, bem como dos fósseis previamente mencionados, e c) dados arqueológicos - artefatos, por exemplo, coletados em diferentes épocas e partes do mundo.
Evidências fósseis
[editar | editar código-fonte]Considerando que os fósseis de Homo sapiens estão restritos apenas àqueles que compartilham uma quantidade significativa de características semelhantes aos humanos existentes nos dias de hoje, nossa espécie, os Homo sapiens, tiveram origem no continente africano, há cerca de 200 mil anos, durante o pleistoceno.[38][39] Chamamos os Homo sapiens de humanos anatomicamente modernos, isto é, humanos que tinham características físicas semelhantes às que observamos nas populações humanas atualmente.[3][40] No entanto, há a denominação “anatomicamente modernos” porque em algumas situações é difícil determinar como era o comportamento, ou seja, quais eram as habilidades cognitivas e sociais daquele indivíduo ou população. Os registros arqueológicos são muitos valiosos se tratando dessa questão, e apresentaremos algumas contribuições da arqueologia mais abaixo. Aqui, faremos um recorte dentro do vasto estudo paleológico, apresentando fósseis que constituíram peças chave para o entendimento da história da evolução humana.
Os Homo sapiens tiveram origem a partir de uma longa linhagem de hominídeos, tendo como ancestral imediato os Homo heidelbergensis, espécie que surgiu há cerca de 0,7 milhões de anos. Na Europa e na Ásia, o Homo heidelbergensis também originou os Neandertais e Denisovanos, respectivamente. Dessa forma, o Homo heidelbergensis é nosso ancestral comum com Neandertais e Denisovanos.[41]
Apesar do surgimento dos primeiros Homo sapiens modernos ser estimado em 200 mil anos antes do presente, é difícil determinar o momento exato em que uma espécie surge. Assim, na comunidade científica existem discussões acerca de como se deu - e quão rápido foi - o surgimento do Homo sapiens anatomicamente moderno, a partir dos humanos arcaicos. Uma forma de elucidar essa questão é através da descoberta de fósseis com características intermediárias. Essa é a natureza dos fósseis encontrados em Jebel Irhoud, Marrocos. Esses fósseis datam entre 300 e 350 mil anos de idade e fazem parte do grupo de fósseis mais antigos do Homo sapiens moderno, tendo ainda características mais antigas em relação à aparência dos humanos nos dias de hoje. Dessa forma, esses fósseis marcam a transição de traços arcaicos - por exemplo, dentes maiores, testa mais proeminente e menor cavidade cerebral - para as características físicas atualmente observadas nos humanos.[39]
Essas informações não implicam no Homo sapiens ter surgido em Marrocos - em oposição aos fósseis de Omo Kibish, na Etiópia, até então os fósseis mais antigos encontrados de humanos modernos - mas ilustra como o Homo sapiens teve, possivelmente, uma origem pan-africana, ou seja, que o processo evolutivo do Homo sapiens envolveu diversas partes do continente africano.[39] Por sua vez, os fósseis de Omo Kibish, denominados Omo Kibish 1 e Omo Kibish 2, são fósseis datados como tendo aproximadamente 200 mil anos e marcam o início do Homo sapiens anatomicamente moderno, dado que a natureza dos fósseis de Jebel Irhoud segue sendo debatida.[39][3]
Existe uma série de fósseis datados de épocas posteriores aos fósseis mencionados acima. Seguindo a cronologia da evolução humana, tudo indica que os humanos modernos surgiram na África.[3] Um estudo de 2018 sugere que o primeiro registro de humanos anatomicamente modernos fora do continente africano esteja na região do Levante.[42] Os sítios Es-Skhul e Qafzeh, por muito tempo, foram conhecidos por abrigar os fósseis mais antigos do Homo sapiens moderno fora do continente africano, datados de 90 a 125 mil anos. A hipótese defendia que os fósseis eram evidências das primeiras tentativas de dispersão humana da África tropical para a região temperada da Eurásia.[3] Com estudos posteriores, foi possível compreender que as migrações mais antigas podem ter resultado em extinção, por diversos motivos, como possíveis competições e outras interações com espécies humanas já extintas, por exemplo os Neandertais.[3]
Descobertas mais recentes na caverna Misliya, em Israel, indicam que essa primeira dispersão para fora da África tenha acontecido há 180 mil anos, muito antes do que era esperado até então.[42] As evidências fósseis sugerem que os humanos modernos interagiram com outras espécies de humanos já extintas, por exemplo os neandertais, por milhares de anos. Contudo, as novas datações fósseis indicam, ainda, que podem ser encontrados fósseis de humanos modernos ainda mais antigos no oeste asiático. Descobertas em Daoxian e Zhirendong, na China, por exemplo, indicam ondas de migração à Europa e Ásia mais antigas do que as descobertas no Levante, região do Oriente Médio.[3]
Muito embora a importância das análises morfológicas em estudos de registros fósseis seja evidente para a compreensão da origem e dispersão dos humanos, a paleoantropologia ainda apresenta limitações e passa por mudanças que permitem aprimorar pesquisas e descobertas na área. Desde 1950, os estudos desse ramo passaram por falta de confiabilidade em métodos de datação de restos mortais, cronologicamente organizados de acordo com as suposições de cada pesquisador. Outro obstáculo está relacionado à falta de uma abordagem sistemática para a análise da morfologia esquelética, bem como a seleção e medição subjetiva dos fósseis, que ao longo da história se misturaram com questões de preconceito racial generalizado na abordagem da evolução humana. Por último a qualidade de preservação e a fragmentação das peças.[3]
Por meio de novos métodos e técnicas de estudo, a paleoantropologia está, aos poucos, passando por reestruturações e revalidação de muitas evidências fósseis já estudadas. A integração de várias disciplinas, principalmente com o auxílio de instrumentos da biologia molecular permite construir filogenias com maior assertividade. Ademais, o aprimoramento de datações radiométricas e técnicas avançadas para análise dos fósseis contribui para maior confiabilidade dos resultados encontrados.[3]
.jpg)

| Sítio | Idade Estimada
(Método de datação) |
Capacidade
Craniana (cc) |
Características arcaicas |
|---|---|---|---|
| Djebel Irhoud
(Marrocos) |
300,000 anos (TL, ESR,
U-series, Bioestratigrafia) |
1,305 e
1,450 |
Caixa craniana alongada, arcada supraciliar contínua |
| Guomde
(Quénia) |
270,000 anos (ESR,
U-series, Estratigrafia) |
- | Arcada supraciliar contínua |
| Florisbad | 300,000-150,000 anos
(ESR, Bioestratigrafia) |
- | Paredes craniais espessas, face com leve prognatismo |
| Eliye Springs
(Quénia) |
Mais de 200,000 anos
(Estratigrafia) |
Ca. 1,210 | Baixa capacidade craniana, abóbada craniana baixa,
ângulo occipital na faixa de Homo erectus |
| Omo-Kibish
(Etiópia) |
195,000-104,000 anos
(U-series, argon-argon) |
1,435
(Omo II) |
Crista sagital, osso occipital angulado |
| Herto (Etiópia) | 160,000 anos
(argon-argon) |
1,450 | Osso occipital angulado |
| Singa (Sudão) | 170,000-150,000 anos
(U/Th, ESR, Bioestratigrafia) |
1,550 | Osso frontal altamente convexo,
arcada supraciliar espessa |
| Laetoli
(Tanzânia) |
120,000 anos
(U-series, Bioestratigrafia) |
1,367
(Laetoli 18) |
Escama do osso frontal plana e estreita, lateral
da arcada supraciliar espessa e arredondada |
| Cavernas Rio
Klassies (África do Sul) |
115,000-60,000 anos
(ESR, Bioestratigrafia) |
- | Osso zigomático grande e robusto, dentes pequenos,
processo coronóide da ulna baixo |
| Border Cave
(África do Sul) |
170,000-50,000 anos
(U-series, ESR, casca ovo de avestruz) |
1,510 | - |
| Dar es-Soltan
II (Marrocos) |
128,000-40,000 anos
(Estratigrafia Arqueológica) |
- | Arcada supraciliar presente em um indivíduo |
| Taramsa Hill
(Egito) |
80,000-50,000 years
(OSL) |
- | Anatomia supraorbital e neurocraniana
"quase modernas" |
| Caverna Blombos
(África do Sul) |
Ca. 75,000 anos (OSL) | - | - |
Arqueologia
[editar | editar código-fonte]Artefatos, isto é, objetos ou indícios de objetos que ajudam a relatar como as sociedades humanas viviam no passado, são parte do registro arqueológico e ajudam a entender como se deu a dispersão humana e quais foram suas consequências. Artefatos também podem nos ajudar a compreender aspectos comportamentais. O comportamento humano moderno é geralmente resumido em habilidades cognitivas e instrumentais sofisticadas, organização social e pensamento simbólico.[50]
Nosso ancestral comum com o Homo neanderthalensis, o Homo heidelbergensis, já produzia ferramentas e armas considerados complexos, na medida em que eram compostos por ao menos três componentes, como a confecção de lanças. A criação de artefatos complexos é um indicativo de uma transição do gênero Homo em relação à capacidade de manipular, armazenar e transmitir informações.[3] Acredita-se que os dados arqueológicos também fornecem informações sobre intercâmbio cultural entre Neandertais e homens modernos. Um exemplo seria a indústria châtelperroniana da Região da Cantábria. Nela foram encontrados ferramentas de pedra caracteristicamente produzido por Neandertais e outros artefatos diagnosticamente produzidos por homens modernos.[3]
Novas categorias de artefatos e informações não genéticas surgiram de forma gradual e sequencial com as migrações do Homo sapiens, indicando possível padrão de desenvolvimento e complexidade da cognição humana moderna. Entre elas estão os ornamentos como objetos para exibição material, a arte visual como esculturas, objetos com notações e instrumentos musicais que apresentam registros apenas posteriores a 50 000 anos antes do presente. Já objetos utilizados para ornamento pessoal, por exemplo conchas marinhas perfuradas, por exemplo, datam de há 135 000 anos na região do Norte da África. Algumas hipóteses sugerem, entretanto, que alguns desses objetos, em especial os de exibição social, sejam evolução de formas anteriores construídas por hominínios primitivos.[51]
Há 75 mil anos, quando os humanos modernos já se dispersavam a partir da África para outros continentes, os registros indicam que os humanos adquiriram a habilidade de pensamento complexo. Assim, os artefatos encontrados que datam de 75 mil anos na África apresentam complexidades associadas às características cognitivas dos humanos modernos e complementam o estudo da anatomia dos fósseis encontrados para dar força à hipótese de que os humanos modernos surgiram na África[3]
Entre os eventos mais marcantes do desenvolvimento cognitivo do Homo sapiens estão o surgimento repentino das artes visuais e de notações simbólicas que não aparecem em outras espécies. A capacidade humana de traduzir informações do cérebro em uma estrutura, a linguagem sintática e a artefatos complexos, incluindo autômatos. Outras formas de informação como a escrita e a eletrônica parecem ter surgido após a era glacial. Já as pinturas e decorações corporais com desenhos feitos à base de pigmento natural, carvão e argila têm datação de mais de 60.000 anos atrás.
Artefatos de maior complexidade - por exemplo, armadilhas, armas de caça, objetos utilizados para costura ou para cozinhar, objetos possivelmente simbólicos ou religiosos - em relação aos objetos mais antigos feitos por hominídeos aparecem no registro arqueológico de maneira que coincidem com as hipóteses de dispersão dos humanos modernos pela África e para além do continente africano. A existência desses artefatos pode ser registrada de maneira direta, ou seja, a presença de uma agulha indica que aquela população de humanos costurava, ou indireta, como: a presença de ômega 3 nos fósseis humanos indica que a sociedade em que aquele indivíduo viveu tinha algum tipo de tecnologia de pesca - varas de pesca, por exemplo.[52] Exemplos de artefatos complexos construídos por humanos modernos são as primeiras máquinas de ação automática, os autômatos, como armadilhas feitas para capturar pequenos mamíferos. Entre as primeiras evidências de autómatos, pode-se citar os restos faunísticos da Caverna de Sibudu (KwaZulu-Natal), na África do Sul, datando de cerca de 65.000 a 62.000 anos atrás, que sugere o possível uso de armadilhas.[53]
Apesar de ajudar a compreender a evolução da capacidade cognitiva e da dispersão do Homo sapiens, a atribuição de muitos vestígios arqueológicos a humanos modernos ou não modernos passa por incertezas em várias partes do mundo. Além disso, alguns materiais utilizados para a fabricação de artefatos não são resistentes e se deterioram ao longo do tempo. As raras evidências de artes visuais datadas de 75.000 anos atrás eram feitas de madeira, indicando que muito material pode ter sido perdido.[3]
Genética
[editar | editar código-fonte]
O campo das evidências genéticas vem se desenvolvendo à medida que se torna mais fácil analisar o DNA. No século XXI, passaram a existir técnicas que tornam viável o sequenciamento do DNA completo de um organismo. Assim, a quantidade de dados genéticos aumentou muito, e agora as evidências genéticas compõem uma parte muito importante da discussão sobre a origem dos humanos modernos.[3]
DNA Mitocondrial
[editar | editar código-fonte]Não só o genoma do indivíduo é importante ao inferir ancestralidade - uma das estratégias mais sólidas para simulação de filogenias se concentra na análise do DNA mitocondrial (mtDNA). A mitocôndria tem um trecho de bases nitrogenadas que formam a estrutura do seu DNA circular - que não sofre recombinação - e grande parte dessa sequência não participa no processo de síntese de proteínas. Por não sofrer pressão seletiva, portanto, está muito suscetível a acumular mutações, ocasionadas por pequenos erros durante a duplicação do DNA que, por sua vez, ocorre durante o processo de divisão celular. Por conta de cada célula possuir muitas mitocôndrias, há grande abundância de DNA mitocondrial em poucas porções de tecido. Além disso, em humanos, mitocôndrias são transmitidas somente das mães para os filhos - assim, inferências filogenéticas podem ter maior resolução em menor espaço de tempo e se tornam mais confiáveis.[54]

Em 1972 Richard Lewontin apontou baixa variabilidade no mtDNA de mulheres desde a dispersão global de humanos modernos. Por consequência da análise da árvore filogenética construída a partir desses dados, concluíram que mulheres modernas tiveram origem de uma ancestral que viveu há aproximadamente 200 mil anos, na África. Na mídia da época, a repercussão fez com que essa mulher fosse batizada de Eva Mitocondrial.[55][56][57]
Cromossomo Y
[editar | editar código-fonte]Assim como o DNA mitocondrial, o cromossomo Y (Y-DNA) não sofre recombinação, mas tem um número muito maior de pares de bases em comparação com o exemplo anterior (aproximadamente 58 milhões; mDNA tem aproximadamente 16500). É a contraposição às linhagens baseadas em mtDNA, pelo cromossomo Y ser passado exclusivamente a homens. Um estudo recriou uma filogenia que estima que a origem do ancestral africano masculino data de apenas 59 mil anos, o que foi complementado com evidências fósseis da caverna Feldhofer na Alemanha - onde encontraram o “Neandertal original”.
Genômica
[editar | editar código-fonte]As primeiras análises genômicas com representação de toda população mundial renderam resultados que já eram previstos pela hipótese da Origem Africana Recente, tendo sido observada maior diversidade genética entre as populações africanas e uma diversidade decrescente conforme se afasta da África ao longo das rotas de migrações dos antepassados que fundaram as diferentes populações ao redor do mundo (devido principal aos efeito de gargalo populacional e efeito fundador, como previsto pela genética populacional).[58][33] Um estudo que analisou microssatélites em todo o genoma de 3000 indivíduos africanos, encontrou 14 agrupamentos populacionais ancestrais que estão fortemente correlacionados aos padrões geográficos, culturais e linguísticos observados por toda a África.[59]
Em 2016, três estudos independentes publicados na revista Nature, utilizando dados de indivíduos de mais de 270 localidades pelo mundo, obtidos através de técnicas modernas de alta qualidade de sequenciamento de todo o genoma, apontaram para a ocorrência de uma uma única grande dispersão a partir da África, a partir da qual todos os não-africanos viventes derivaram de uma mesma população ancestral advinda da África,[60][61][62] com uma possível contribuição de uma migração anterior à Oceania.[62]
Outras evidências
[editar | editar código-fonte]Além de evidências genéticas, fósseis e arqueológicas, existem outras evidências que sustentam a hipótese da origem única. Estudos recentes demonstram que das três grandes famílias de bactérias intestinais, pelo menos duas delas coevoluiram com a espécie humana há milhões de anos.[63] Pesquisadores sugerem que esta relação íntima entre o homem e a bactéria intestinal Helicobacter pylori data antes mesmo do início das ondas migratórias do homem para o resto do mundo.[39] Por conta disso, na última década, através de análises moleculares do DNA da H. pylori obtido de várias pessoas de lugares diferentes do mundo, foi demonstrado que a diversidade genética deste microrganismo reflete a imigração humana e, posteriormente, as separações geográficas e étnicas entre os grupos humanos. Por exemplo, em um estudo realizado, pesquisadores compararam sequências específicas do DNA de H. pylori extraídos de hospedeiros das regiões de Connecticut e Malawi, foi descoberto que as cepas bacterianas de Malawi divergiram da cepa americana há, pelo menos, 1.7 milhões de anos, que corresponde, exatamente, à diáspora mais recente do homem saindo da África. Isso ocorre porque as sequências de fragmentos de sete genes de manutenção e um gene associado à virulência (vacA) diferem de acordo com o continente de origem do hospedeiro, podendo, assim, dividir a bactéria em sete populações e subpopulações com distribuições geográficas diferentes entre si.[64] Além disso, outros estudos sugerem que, assim como ocorre com os humanos, a diversidade genética de H. pylori diminui conforme aumenta a distância do Leste da África, o berço do homem moderno.[39] E a análise comparativa do DNA dos piolhos do corpo humano (Pediculus humanus), que habitam as roupas, também revela uma origem africana recente (data de dispersão estimada em cerca de 72.000 anos atrás).[3]
Também, evidências linguísticas sugerem que fonemas (unidades de som) presentes nas línguas modernas exibe o mesmo padrão observado no DNA, ou seja, a diversidade diminui em função do aumento da distância da África, sendo este fenômeno explicado pelo efeito fundador, em que sucessivos gargalos populacionais ocorridos durante a expansão seriam responsáveis por reduzir a diversidade de forma progressiva.[65]
Dados climatológicos também são usados como evidências para a hipótese única, uma vez que dados fósseis e climatológicos de oito milhões de anos atrás sugerem que alguns eventos ocorridos durante a evolução da espécie humana, tais como, evolução de novas espécies, dispersão da África para outros continentes, correspondem a períodos de mudanças climáticas na África e Eurásia. Por exemplo, cerca de 38.000 a 36.000 anos atrás, houve um período de calor intenso, responsável por uma explosão demográfica da espécie humana na região norte da Eurásia.[66]
Hibridação
[editar | editar código-fonte]
Desde 2010 vem se acumulando evidências de hibridação entre Homo sapiens e outras linhagens hominínias, como H. neanderthalensis e H. denisova, através de estudos de análise genômica, que, pela quantidade muito maior de dados gerada, permite se acessar com maior confiabilidade as relações filogenéticas entre as espécies, reconstruindo os genomas das espécies hominínias extintas.[33][36][35]
As espécies H. neanderthalensis e H. denisova divergiram de H. sapiens a partir de um ancestral comum (provavelmente representado por H. heidelbergensis ou H. rhodesiensis) que migrou da África a aproximadamente 500 mil anos atrás, dando origem aos Neandertais na Europa e Denisovanos na Ásia. Quando os Homo sapiens modernos se expandiram pelo mundo após 75 mil anos atrás encontraram esses outros hominínios, o que resultou em eventos de hibridação entre essas espécies.[3]
As análises genômicas mais recentes apontam que todas as populações não-africanas possuem cerca de 2% de ancestralidade Neandertal[67][68][69] (possivelmente indicando um evento de hibridação significativo pouco após a principal onda migratória para fora da África).[33] Através dos padrões de desequilíbrio de ligação genética Sriram Sankararaman e pares estimaram a data de hibridação entre humanos e Neandertais para aproximadamente 50–65 mil anos atrás.[70] Porém outros trabalhos apontam para um padrão mais complexo para essa mistura, com diferentes eventos de hibridação em diferentes localidades e momentos, desde a chegada de H. sapiens Eurasia, até a extinção dessas outras espécies de hominínios.[71]
As tecnicas de sequenciamento e análise de genomas também permitiram identificar introgressão de material genético do outra espécie de hominínio, Homo denisova, pouco após seu descobrimento a partir de um fragmento de osso do dedo encontrado na Sibéria.[36] Hoje se estima que populações nativas da Oceania possuem entre 3-6% [36] e populações do leste asiático 0,1-0,3% de material genético possível de ser traçado aos Denisovanos.[72][33]
Uma análise do genoma de 1523 indivíduos de diferentes regiões do mundo identificou que ocorreram ao menos 4 pulsos de fluxo genético entre H. Sapiens e essas duas espécies: (1) um evento pouco após a saída dos humanos modernos da África, contribuindo com uma pequena porcentagem do material genético Neandertal em não-africanos; (2) posterior hibridação com Neandertais contribuindo com a herança genética das populações viventes europeias, sul e leste asiáticas; (3) seguida por cruzamento entre asiáticos do leste e Neandertais; e somente posteriormente tendo ocorrido uma hibridação com Denisovanos que hoje é representada pela porção atribuível a Denisovanos que populações nativas da Oceania (Melanésia e Austrália) carregam.[73]
Outra linha de evidência para a mistura entre humanos modernos e Neandertais está na análise genética de microorganismos e comparação entre as diferentes linhagens encontradas, como de uma bactéria comensal da microbiota oral recuperada do cálculo dental de um fóssil Neandertal[74] e do vírus sexualmente transmitido HPV 16.[75]
Também foram identificadas, a partir de análises do genoma de diversas populações africanas viventes, sinais de hibridação limitada entre H. sapiens moderno e outros representantes "arcaicos" do gênero Homo.[76][77] Com evidências de uma introgressão de aproximadamente 2% tão cedo quanto há 35 mil anos de uma população que divergiu da linhagem humana há aproximadamente 700 mil anos.[78]
Hoje o debate gira em torno de qual é o grau dessa mistura entre H. sapiens e outras espécies do gênero Homo, quanto isso teria influenciado o caminho evolutivo de nossa espécie e de que forma isso se adequa aos diferentes modelos para a origem humana, sendo uma posição bem balanceada face às novas evidências se apontar para um modelo que mantém a Origem Africana Recente como principal contribuidora das características humanas modernas e se adiciona o que pode ser chamado de "Modelo da Origem Africana Recente + Hibridação"[6] ou "Modelo da Substituição com vazamento".[33]
Ver também
[editar | editar código-fonte]Referências
- ↑ Rieux, Adrien; Eriksson, Anders; Li, Mingkun; Sobkowiak, Benjamin; Weinert, Lucy A.; Warmuth, Vera; Ruiz-Linares, Andres; Manica, Andrea; Balloux, François (5 de agosto de 2014). «Improved Calibration of the Human Mitochondrial Clock Using Ancient Genomes». Molecular Biology and Evolution (10): 2780–2792. ISSN 1537-1719. doi:10.1093/molbev/msu222. Consultado em 20 de julho de 2021
- ↑ Poznik, G David; Xue, Yali; Mendez, Fernando L; Willems, Thomas F; Massaia, Andrea; Wilson Sayres, Melissa A; Ayub, Qasim; McCarthy, Shane A; Narechania, Apurva (25 de abril de 2016). «Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences». Nature Genetics (6): 593–599. ISSN 1061-4036. doi:10.1038/ng.3559. Consultado em 19 de julho de 2021
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z Hoffecker, John F. (2017). Modern humans : their African origin and global dispersal. New York: [s.n.] OCLC 988087435
- ↑ Stringer, C.; Andrews, P (2 de março de 1988). «Genetic and fossil evidence for the origin of modern humans». Science (em inglês) (4845): 1263–1268. ISSN 0036-8075. doi:10.1126/science.3125610. Consultado em 25 de junho de 2021
- ↑ Bae, Christopher J.; Douka, Katerina; Petraglia, Michael D. (8 de dezembro de 2017). «On the origin of modern humans: Asian perspectives». Science (em inglês) (6368): eaai9067. ISSN 0036-8075. doi:10.1126/science.aai9067. Consultado em 25 de junho de 2021
- ↑ a b c Stringer, Chris (maio de 2014). «Why we are not all multiregionalists now». Trends in Ecology & Evolution (5): 248–251. ISSN 0169-5347. doi:10.1016/j.tree.2014.03.001. Consultado em 19 de julho de 2021
- ↑ a b c d Stringer, Chris (maio de 2012). «What makes a modern human». Nature (7396): 33–35. ISSN 0028-0836. doi:10.1038/485033a. Consultado em 14 de julho de 2021
- ↑ Stringer, Chris; Galway-Witham, Julia (8 de junho de 2017). «On the origin of our species». Nature (em inglês) (7657): 212–214. ISSN 0028-0836. doi:10.1038/546212a. Consultado em 14 de julho de 2021
- ↑ Bae, Christopher J.; Douka, Katerina; Petraglia, Michael D. (8 de dezembro de 2017). «On the origin of modern humans: Asian perspectives». Science (em inglês) (6368). ISSN 0036-8075. PMID 29217544. doi:10.1126/science.aai9067. Consultado em 14 de julho de 2021
- ↑ a b c d Tattersall, I. (22 de setembro de 2009). «Human origins: Out of Africa». Proceedings of the National Academy of Sciences (em inglês) (38): 16018–16021. ISSN 0027-8424. PMC 2752574
 . PMID 19805256. doi:10.1073/pnas.0903207106. Consultado em 16 de julho de 2021
. PMID 19805256. doi:10.1073/pnas.0903207106. Consultado em 16 de julho de 2021
- ↑ a b Klein, Richard G. (junho de 1995). «Anatomy, behavior, and modern human origins». Journal of World Prehistory (2): 167–198. ISSN 0892-7537. doi:10.1007/bf02221838. Consultado em 17 de julho de 2021
- ↑ a b Mcbrearty, Sally; Brooks, Alison S. (novembro de 2000). «The revolution that wasn't: a new interpretation of the origin of modern human behavior». Journal of Human Evolution (5): 453–563. ISSN 0047-2484. doi:10.1006/jhev.2000.0435. Consultado em 17 de julho de 2021
- ↑ a b c Klein, Richard G. (2009). The Human Career. [S.l.]: University of Chicago Press
- ↑ Pearson, Osbjorn M. (22 de fevereiro de 2008). «Statistical and biological definitions of "anatomically modern" humans: Suggestions for a unified approach to modern morphology». Evolutionary Anthropology: Issues, News, and Reviews (1): 38–48. ISSN 1060-1538. doi:10.1002/evan.20155. Consultado em 15 de julho de 2021
- ↑ Tattersall, Ian; Schwartz, Jeffrey H. (22 de fevereiro de 2008). «The morphological distinctiveness of Homo sapiens and its recognition in the fossil record: Clarifying the problem». Evolutionary Anthropology: Issues, News, and Reviews (em inglês) (1): 49–54. doi:10.1002/evan.20153. Consultado em 15 de julho de 2021
- ↑ Klein, Richard G. (junho de 1995). «Anatomy, behavior, and modern human origins». Journal of World Prehistory (2): 167–198. ISSN 0892-7537. doi:10.1007/bf02221838. Consultado em 16 de julho de 2021
- ↑ a b A Reconsideration of the Omo-Kibish Remains and the erectus-sapiens Transition (em inglês). [S.l.]: Routledge. 5 de dezembro de 2016
- ↑ Dean, Christopher; Leakey, Meave G.; Reid, Donald; Schrenk, Friedemann; Schwartz, Gary T.; Stringer, Christopher; Walker, Alan (dezembro de 2001). «Growth processes in teeth distinguish modern humans from Homo erectus and earlier hominins». Nature (6864): 628–631. ISSN 0028-0836. doi:10.1038/414628a. Consultado em 16 de julho de 2021
- ↑ Smith, T. M.; Tafforeau, P.; Reid, D. J.; Grun, R.; Eggins, S.; Boutakiout, M.; Hublin, J.-J. (19 de março de 2007). «Earliest evidence of modern human life history in North African early Homo sapiens». Proceedings of the National Academy of Sciences (15): 6128–6133. ISSN 0027-8424. doi:10.1073/pnas.0700747104. Consultado em 19 de julho de 2021
- ↑ Henshilwood, C. S. (15 de fevereiro de 2002). «Emergence of Modern Human Behavior: Middle Stone Age Engravings from South Africa». Science (5558): 1278–1280. doi:10.1126/science.1067575. Consultado em 17 de julho de 2021
- ↑ Henshilwood, C. (16 de abril de 2004). «Middle Stone Age Shell Beads from South Africa». Science (em inglês) (5669): 404–404. ISSN 0036-8075. doi:10.1126/science.1095905. Consultado em 17 de julho de 2021
- ↑ Yellen, J.; Brooks, A.; Cornelissen, E; Mehlman, M.; Stewart, K (28 de abril de 1995). «A middle stone age worked bone industry from Katanda, Upper Semliki Valley, Zaire». Science (em inglês) (5210): 553–556. ISSN 0036-8075. doi:10.1126/science.7725100. Consultado em 17 de julho de 2021
- ↑ Kennedy, Kenneth A. R. (dezembro de 1983). «Physical:A History of American Physical Anthropology, 1930-1980 . Frank Spencer , ed». American Anthropologist (4): 965–967. ISSN 0002-7294. doi:10.1525/aa.1983.85.4.02a00350. Consultado em 18 de julho de 2021
- ↑ Robbins, Lawrence H.; Coppens, Y. (abril de 1977). «Earliest Man and Environment in the Lake Rudolf Basin: Stratigraphy, Paleoecology and Evolution.». Man (1). 185 páginas. ISSN 0025-1496. doi:10.2307/2801014. Consultado em 18 de julho de 2021
- ↑ Mellars, Paul (8 de janeiro de 2004). «Archaeology and the Origins of Modern Humans: European and African Perspectives». British Academy. ISBN 978-0-19-726311-2. Consultado em 18 de julho de 2021
- ↑ Cann, Rebecca L.; Stoneking, Mark; Wilson, Allan C. (janeiro de 1987). «Mitochondrial DNA and human evolution». Nature (em inglês) (6099): 31–36. ISSN 0028-0836. doi:10.1038/325031a0. Consultado em 18 de julho de 2021
- ↑ Hoffecker, John F. (2017). Modern humans : their African origin and global dispersal. New York: [s.n.] OCLC 988087435
- ↑ Pääbo, Svante; Poinar, Hendrik; Serre, David; Jaenicke-Després, Viviane; Hebler, Juliane; Rohland, Nadin; Kuch, Melanie; Krause, Johannes; Vigilant, Linda (29 de novembro de 2004). «Genetic Analyses from Ancient DNA». Annual Review of Genetics (1): 645–679. ISSN 0066-4197. doi:10.1146/annurev.genet.37.110801.143214. Consultado em 18 de julho de 2021
- ↑ «Neandertal DNA sequences and the origin of modern humans». Trends in Genetics (10). 391 páginas. Outubro de 1997. ISSN 0168-9525. doi:10.1016/s0168-9525(97)90010-x. Consultado em 18 de julho de 2021
- ↑ Underhill, Peter A.; Shen, Peidong; Lin, Alice A.; Jin, Li; Passarino, Giuseppe; Yang, Wei H.; Kauffman, Erin; Bonné-Tamir, Batsheva; Bertranpetit, Jaume (novembro de 2000). «Y chromosome sequence variation and the history of human populations». Nature Genetics (3): 358–361. ISSN 1061-4036. doi:10.1038/81685. Consultado em 18 de julho de 2021
- ↑ Li, J. Z.; Absher, D. M.; Tang, H.; Southwick, A. M.; Casto, A. M.; Ramachandran, S.; Cann, H. M.; Barsh, G. S.; Feldman, M. (22 de fevereiro de 2008). «Worldwide Human Relationships Inferred from Genome-Wide Patterns of Variation». Science (em inglês) (5866): 1100–1104. ISSN 0036-8075. doi:10.1126/science.1153717. Consultado em 18 de julho de 2021
- ↑ Henn, B. M.; Gignoux, C. R.; Jobin, M.; Granka, J. M.; Macpherson, J. M.; Kidd, J. M.; Rodriguez-Botigue, L.; Ramachandran, S.; Hon, L. (29 de março de 2011). «Hunter-gatherer genomic diversity suggests a southern African origin for modern humans». Proceedings of the National Academy of Sciences (em inglês) (13): 5154–5162. ISSN 0027-8424. PMC 3069156. PMID 21383195. doi:10.1073/pnas.1017511108. Consultado em 18 de julho de 2021
- ↑ a b c d e f Nielsen, Rasmus; Akey, Joshua M.; Jakobsson, Mattias; Pritchard, Jonathan K.; Tishkoff, Sarah; Willerslev, Eske (janeiro de 2017). «Tracing the peopling of the world through genomics». Nature (em inglês) (7637): 302–310. ISSN 1476-4687. PMC 5772775. PMID 28102248. doi:10.1038/nature21347. Consultado em 19 de julho de 2021
- ↑ Trinkaus, E.; Moldovan, O.; Milota, s.; Bilgar, A.; Sarcina, L.; Athreya, S.; Bailey, S. E.; Rodrigo, R.; Mircea, G. (22 de setembro de 2003). «An early modern human from the Pestera cu Oase, Romania». Proceedings of the National Academy of Sciences (20): 11231–11236. ISSN 0027-8424. doi:10.1073/pnas.2035108100. Consultado em 20 de julho de 2021
- ↑ a b Green, R. E.; Krause, J.; Briggs, A. W.; Maricic, T.; Stenzel, U.; Kircher, M.; Patterson, N.; Li, H.; Zhai, W. (7 de maio de 2010). «A Draft Sequence of the Neandertal Genome». Science (em inglês) (5979): 710–722. ISSN 0036-8075. PMC 5100745. PMID 20448178. doi:10.1126/science.1188021. Consultado em 19 de julho de 2021
- ↑ a b c d Reich, David; Green, Richard E.; Kircher, Martin; Krause, Johannes; Patterson, Nick; Durand, Eric Y.; Viola, Bence; Briggs, Adrian W.; Stenzel, Udo (dezembro de 2010). «Genetic history of an archaic hominin group from Denisova Cave in Siberia». Nature (em inglês) (7327): 1053–1060. ISSN 0028-0836. PMC 4306417. PMID 21179161. doi:10.1038/nature09710. Consultado em 19 de julho de 2021
- ↑ Stringer, Chris (maio de 2014). «Why we are not all multiregionalists now». Trends in Ecology & Evolution (em inglês) (5): 248–251. doi:10.1016/j.tree.2014.03.001. Consultado em 19 de julho de 2021
- ↑ Stringer, Chris (5 de julho de 2016). «The origin and evolution of Homo sapiens». Philosophical Transactions of the Royal Society B: Biological Sciences (1698). 20150237 páginas. PMC 4920294. PMID 27298468. doi:10.1098/rstb.2015.0237. Consultado em 20 de julho de 2021
- ↑ a b c d e f Hublin, Jean-Jacques; Ben-Ncer, Abdelouahed; Bailey, Shara E.; Freidline, Sarah E.; Neubauer, Simon; Skinner, Matthew M.; Bergmann, Inga; Le Cabec, Adeline; Benazzi, Stefano (junho de 2017). «New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens». Nature (em inglês) (7657): 289–292. ISSN 1476-4687. doi:10.1038/nature22336. Consultado em 20 de julho de 2021
- ↑ Pearson, Osbjorn M. (2008). «Statistical and biological definitions of "anatomically modern" humans: Suggestions for a unified approach to modern morphology». Evolutionary Anthropology: Issues, News, and Reviews (em inglês) (1): 38–48. ISSN 1520-6505. doi:10.1002/evan.20155. Consultado em 20 de julho de 2021
- ↑ Buck, Laura T.; Stringer, Chris B. (março de 2014). «Homo heidelbergensis». Current Biology (6): R214–R215. ISSN 0960-9822. doi:10.1016/j.cub.2013.12.048. Consultado em 20 de julho de 2021
- ↑ a b Callaway, Ewen (25 de janeiro de 2018). «Israeli fossils are the oldest modern humans ever found outside of Africa». Nature (em inglês) (7690): 15–16. doi:10.1038/d41586-018-01261-5. Consultado em 20 de julho de 2021
- ↑ Trinkaus, Erik (outubro de 2005). «Early Modern Humans». Annual Review of Anthropology (1): 207–230. ISSN 0084-6570. doi:10.1146/annurev.anthro.34.030905.154913. Consultado em 14 de julho de 2021
- ↑ Bräuer, Günter (22 de fevereiro de 2008). «The origin of modern anatomy: By speciation or intraspecific evolution?». Evolutionary Anthropology: Issues, News, and Reviews (1): 22–37. ISSN 1060-1538. doi:10.1002/evan.20157. Consultado em 14 de julho de 2021
- ↑ Mellars, Paul (8 de janeiro de 2004). «Archaeology and the Origins of Modern Humans: European and African Perspectives». British Academy. ISBN 978-0-19-726311-2. Consultado em 14 de julho de 2021
- ↑ author., Klein, Richard G.,. The human career : human biological and cultural origins. [S.l.: s.n.] OCLC 233283760
- ↑ Lieberman, Daniel E. (3 de maio de 2011). The Evolution of the Human Head. [S.l.]: Harvard University Press
- ↑ Hublin, Jean-Jacques; Ben-Ncer, Abdelouahed; Bailey, Shara E.; Freidline, Sarah E.; Neubauer, Simon; Skinner, Matthew M.; Bergmann, Inga; Le Cabec, Adeline; Benazzi, Stefano (junho de 2018). «Author Correction: New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens». Nature (7711): E6–E6. ISSN 0028-0836. doi:10.1038/s41586-018-0166-3. Consultado em 14 de julho de 2021
- ↑ Richter, Daniel; Grün, Rainer; Joannes-Boyau, Renaud; Steele, Teresa E.; Amani, Fethi; Rué, Mathieu; Fernandes, Paul; Raynal, Jean-Paul; Geraads, Denis (8 de junho de 2017). «The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age». Nature (7657): 293–296. ISSN 0028-0836. doi:10.1038/nature22335. Consultado em 14 de julho de 2021
- ↑ Tylén, Kristian, et al. "The evolution of early symbolic behavior in Homo sapiens." Proceedings of the National Academy of Sciences 117.9 (2020): 4578-4584.
- ↑ Marek Kohn and Steven J. Mithen, “Handaxes: Products of Sexual Selection?” Antiquity 73 (1999): 518–526; April Nowell and Melanie Lee Chang, “The Case Against Sexual Selection as an Explanation of Handaxe Morphology,” Paleo-Anthropology (2009): 77–88.
- ↑ Hoffecker, John F. (2017). Modern humans : their African origin and global dispersal. New York: [s.n.] OCLC 988087435.
- ↑ Lyn Wadley, “Were Snares and Traps Used in the Middle Stone Age and Does It Matter? A Review and a Case Study from Sibudu, South Africa,” Journal of Human Evolution 58 (2020): 179–192.
- ↑ Cann, Rebecca L.; Stoneking, Mark; Wilson, Allan C. (janeiro de 1987). «Mitochondrial DNA and human evolution». Nature (6099): 31–36. ISSN 0028-0836. doi:10.1038/325031a0. Consultado em 20 de julho de 2021
- ↑ Lewontin, R. C. (1972). «The Apportionment of Human Diversity». New York, NY: Springer US: 381–398. ISBN 978-1-4684-9065-7. Consultado em 20 de julho de 2021
- ↑ Templeton, A. (7 de fevereiro de 1992). «Human origins and analysis of mitochondrial DNA sequences». Science (5045): 737–737. ISSN 0036-8075. doi:10.1126/science.1590849. Consultado em 20 de julho de 2021
- ↑ Horai, S.; Hayasaka, K.; Kondo, R.; Tsugane, K.; Takahata, N. (17 de janeiro de 1995). «Recent African origin of modern humans revealed by complete sequences of hominoid mitochondrial DNAs.». Proceedings of the National Academy of Sciences (2): 532–536. ISSN 0027-8424. doi:10.1073/pnas.92.2.532. Consultado em 20 de julho de 2021
- ↑ Rosenberg, N. A. (20 de dezembro de 2002). «Genetic Structure of Human Populations». Science (5602): 2381–2385. ISSN 0036-8075. doi:10.1126/science.1078311. Consultado em 22 de julho de 2021
- ↑ Tishkoff, S. A.; Reed, F. A.; Friedlaender, F. R.; Ehret, C.; Ranciaro, A.; Froment, A.; Hirbo, J. B.; Awomoyi, A. A.; Bodo, J.-M. (30 de abril de 2009). «The Genetic Structure and History of Africans and African Americans». Science (5930): 1035–1044. ISSN 0036-8075. doi:10.1126/science.1172257. Consultado em 22 de julho de 2021
- ↑ Malaspinas, Anna-Sapfo; Westaway, Michael C.; Muller, Craig; Sousa, Vitor C.; Lao, Oscar; Alves, Isabel; Bergström, Anders; Athanasiadis, Georgios; Cheng, Jade Y. (13 de outubro de 2016). «A genomic history of Aboriginal Australia». Nature (em inglês) (7624): 207–214. ISSN 0028-0836. doi:10.1038/nature18299. Consultado em 22 de julho de 2021
- ↑ Mallick, Swapan; Li, Heng; Lipson, Mark; Mathieson, Iain; Gymrek, Melissa; Racimo, Fernando; Zhao, Mengyao; Chennagiri, Niru; Nordenfelt, Susanne (13 de outubro de 2016). «The Simons Genome Diversity Project: 300 genomes from 142 diverse populations». Nature (em inglês) (7624): 201–206. ISSN 0028-0836. PMC 5161557. PMID 27654912. doi:10.1038/nature18964. Consultado em 22 de julho de 2021
- ↑ a b Pagani, Luca; Lawson, Daniel John; Jagoda, Evelyn; Mörseburg, Alexander; Eriksson, Anders; Mitt, Mario; Clemente, Florian; Hudjashov, Georgi; DeGiorgio, Michael (13 de outubro de 2016). «Genomic analyses inform on migration events during the peopling of Eurasia». Nature (em inglês) (7624): 238–242. ISSN 0028-0836. PMC 5164938. PMID 27654910. doi:10.1038/nature19792. Consultado em 22 de julho de 2021
- ↑ Correa, Pelayo; Piazuelo, M. Blanca (janeiro de 2012). «Evolutionary History of the Helicobacter pylori Genome: Implications for Gastric Carcinogenesis». Gut and Liver (1): 21–28. ISSN 1976-2283. PMC 3286735. PMID 22375167. doi:10.5009/gnl.2012.6.1.21. Consultado em 20 de julho de 2021
- ↑ Falush, Daniel; Wirth, Thierry; Linz, Bodo; Pritchard, Jonathan K.; Stephens, Matthew; Kidd, Mark; Blaser, Martin J.; Graham, David Y.; Vacher, Sylvie (7 de março de 2003). «Traces of Human Migrations in Helicobacter pylori Populations». Science (em inglês) (5612): 1582–1585. ISSN 0036-8075. PMID 12624269. doi:10.1126/science.1080857. Consultado em 20 de julho de 2021
- ↑ Atkinson, Quentin D. (15 de abril de 2011). «Phonemic Diversity Supports a Serial Founder Effect Model of Language Expansion from Africa». Science (em inglês) (6027): 346–349. ISSN 0036-8075. PMID 21493858. doi:10.1126/science.1199295. Consultado em 20 de julho de 2021
- ↑ Hamilton, Robert M.,; et al. «Understanding Climate's Influence on Human Evolution». The National Academies
- ↑ Wall, Jeffrey D; Yang, Melinda A; Jay, Flora; Kim, Sung K; Durand, Eric Y; Stevison, Laurie S; Gignoux, Christopher; Woerner, August; Hammer, Michael F (1 de maio de 2013). «Higher Levels of Neanderthal Ancestry in East Asians than in Europeans». Genetics (1): 199–209. ISSN 1943-2631. doi:10.1534/genetics.112.148213. Consultado em 20 de julho de 2021
- ↑ Vernot, Benjamin; Akey, Joshua M. (março de 2015). «Complex History of Admixture between Modern Humans and Neandertals». The American Journal of Human Genetics (3): 448–453. ISSN 0002-9297. doi:10.1016/j.ajhg.2015.01.006. Consultado em 20 de julho de 2021
- ↑ Sankararaman, Sriram; Mallick, Swapan; Dannemann, Michael; Prüfer, Kay; Kelso, Janet; Pääbo, Svante; Patterson, Nick; Reich, David (29 de janeiro de 2014). «The genomic landscape of Neanderthal ancestry in present-day humans». Nature (7492): 354–357. ISSN 0028-0836. doi:10.1038/nature12961. Consultado em 20 de julho de 2021
- ↑ Sankararaman, Sriram; Patterson, Nick; Li, Heng; Pääbo, Svante; Reich, David (4 de outubro de 2012). «The Date of Interbreeding between Neandertals and Modern Humans». PLOS Genetics (em inglês) (10): e1002947. ISSN 1553-7404. PMC 3464203. PMID 23055938. doi:10.1371/journal.pgen.1002947. Consultado em 20 de julho de 2021
- ↑ Vernot, Benjamin; Akey, Joshua M. (março de 2015). «Complex History of Admixture between Modern Humans and Neandertals». The American Journal of Human Genetics (em inglês) (3): 448–453. PMC 4375686. PMID 25683119. doi:10.1016/j.ajhg.2015.01.006. Consultado em 20 de julho de 2021
- ↑ Skoglund, P.; Jakobsson, M. (31 de outubro de 2011). «Archaic human ancestry in East Asia». Proceedings of the National Academy of Sciences (45): 18301–18306. ISSN 0027-8424. doi:10.1073/pnas.1108181108. Consultado em 20 de julho de 2021
- ↑ Vernot, B.; Tucci, S.; Kelso, J.; Schraiber, J. G.; Wolf, A. B.; Gittelman, R. M.; Dannemann, M.; Grote, S.; McCoy, R. C. (8 de abril de 2016). «Excavating Neandertal and Denisovan DNA from the genomes of Melanesian individuals». Science (em inglês) (6282): 235–239. ISSN 0036-8075. PMC 6743480. PMID 26989198. doi:10.1126/science.aad9416. Consultado em 20 de julho de 2021
- ↑ Weyrich, Laura S.; Duchene, Sebastian; Soubrier, Julien; Arriola, Luis; Llamas, Bastien; Breen, James; Morris, Alan G.; Alt, Kurt W.; Caramelli, David (abril de 2017). «Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus». Nature (em inglês) (7650): 357–361. ISSN 0028-0836. doi:10.1038/nature21674. Consultado em 20 de julho de 2021
- ↑ Pimenoff, Ville N.; de Oliveira, Cristina Mendes; Bravo, Ignacio G. (janeiro de 2017). «Transmission between Archaic and Modern Human Ancestors during the Evolution of the Oncogenic Human Papillomavirus 16». Molecular Biology and Evolution (em inglês) (1): 4–19. ISSN 0737-4038. PMC 5854117. PMID 28025273. doi:10.1093/molbev/msw214. Consultado em 20 de julho de 2021
- ↑ Hsieh, PingHsun; Woerner, August E.; Wall, Jeffrey D.; Lachance, Joseph; Tishkoff, Sarah A.; Gutenkunst, Ryan N.; Hammer, Michael F. (março de 2016). «Model-based analyses of whole-genome data reveal a complex evolutionary history involving archaic introgression in Central African Pygmies». Genome Research (em inglês) (3): 291–300. ISSN 1088-9051. PMC 4772012. PMID 26888264. doi:10.1101/gr.196634.115. Consultado em 20 de julho de 2021
- ↑ Lachance, Joseph; Vernot, Benjamin; Elbers, Clara C.; Ferwerda, Bart; Froment, Alain; Bodo, Jean-Marie; Lema, Godfrey; Fu, Wenqing; Nyambo, Thomas B. (agosto de 2012). «Evolutionary History and Adaptation from High-Coverage Whole-Genome Sequences of Diverse African Hunter-Gatherers». Cell (em inglês) (3): 457–469. PMC 3426505. PMID 22840920. doi:10.1016/j.cell.2012.07.009. Consultado em 20 de julho de 2021
- ↑ Hammer, M. F.; Woerner, A. E.; Mendez, F. L.; Watkins, J. C.; Wall, J. D. (13 de setembro de 2011). «Genetic evidence for archaic admixture in Africa». Proceedings of the National Academy of Sciences (em inglês) (37): 15123–15128. ISSN 0027-8424. PMC 3174671. PMID 21896735. doi:10.1073/pnas.1109300108. Consultado em 20 de julho de 2021
Fontes
[editar | editar código-fonte]- Modern Men Trace Ancestry to African Migrants, Science, 11 de maio de 2001
- Underhill et al. (2001). "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations" Ann. Hum. Genet. 65: 43-62. PDF. Retrieved 25 July 2007.
- Neanderthals 'mated with modern humans', BBC News, 21 de abril de 1999
- New analysis shows three human migrations out of Africa - Replacement theory 'demolished', Washington University in St. Louis, 2 de fevereiro de 2006
- Long and Kittles (2003). "Human genetic variation and the nonexistence of human races" Human Biology, 75: 449-471. PDF. Retrieved 10 January 2007.
- Risch, N., Burchard, E., Ziv, E. and Tang, H. (2002). «Categorization of humans in biomedical research: genes, race and disease». Genome Biology. 3 (7): comment2007.2001 - comment2007.2012
- Tishkoff, S. and Kidd, K. Implications of biogeography of human populations for 'race' and medicine Nature Genetics 36: S21 - S27 (2004)