
鞭毛
鞭毛(べんもう、英: flagellum、複数形: flagella)は、毛状の細胞小器官で、遊泳に必要な推進力を生み出す事が主な役目である。構造的に真核生物鞭毛と細菌鞭毛、古細菌鞭毛とに分けられる。
真核生物鞭毛
[編集]
動植物の精子から、クラミドモナスやミドリムシといった藻類や原生生物にまで広く見られる。長さは数µmから数十µm、横断面の直径は200-300 nmほどである。通常の明視野型光学顕微鏡でも確認できるが、位相差顕微鏡や暗視野顕微鏡、微分干渉顕微鏡であればより明瞭に観察する事ができる。
動作原理
[編集]真核生物の鞭毛は、鞭毛それ自体が能動的に屈曲し、運動する能力を持つ。鞭毛の断面を電子顕微鏡で観察すると、9+2構造と呼ばれる微小管配置が観察される。鞭毛の中核を成すこの部分を軸糸(axoneme)と呼ぶ。この微小管の間にはダイニンというタンパク質分子モーターが存在する。ダイニンがATPを加水分解してエネルギーを取り出し、これが微小管(A小管とB小管)同士の滑り運動に変換されることで、鞭毛の屈曲が引き起こされると考えられている。
鞭毛の構成要素とタンパク質
[編集]
1A:二連管(A小管)
1B:二連管(B小管)
2:中心対微小管
3:ダイニン外腕
4:スポーク
5:ネキシン
6:細胞膜
鞭毛の表面は細胞膜である。さらにその外側に、種々の修飾構造を持つ生物もいる(後述)。
- 微小管
- 中心微小管(中心対微小管、central pair)
- 周辺微小管(二連管、doublets)
- A小管:やや内側に位置する小管。スポークと接する。
- B小管:やや外側に位置する小管。
微小管は、α-チューブリンとβ-チューブリンが交互に並んで構成された13本の繊維から成る構造である。真核生物特有の構造で、原核生物には存在しない。
- ダイニン(dynein)
- ダイニン外腕
- ダイニン内腕
- ネキシン(ネキシンリンク、nexin)
- スポーク(ラジアルスポーク)
- キネシン(kinesin):モータータンパク質。鞭毛内輸送(IFT; intraflagellar transport)に関与する。
ここに列挙した構造やタンパク質は一例で、実際にはさらに多くの要素を含む。
鞭毛装置(flagellar apparatus)
[編集]鞭毛の根元の基底小体と、それに付随する種々の鞭毛根その他の構造を合わせてこう呼ぶ。生物によっては、パラバサリアの副基体(parabasal body)やハプト藻類のハプトネマなど、鞭毛以外の構造の基部を含む場合もある。基底小体だけで200種以上、鞭毛装置全体では300種以上のタンパク質を含むと言われる。
鞭毛装置は分類群毎の多様性と適度な保存性とを兼ね備える。従って鞭毛装置の形態は、真核生物ほぼ全体の分類に対して通用する、数少ない形態形質である。
- 鞭毛移行帯(transitional region):鞭毛上部(9+2構造の部分)と基底小体の間の領域。鞭毛装置には含めない事もある。この部位の機能は定かでないが、分類群によっては独自の構造を持つ。
- 基底小体(basal body)
- 鞭毛の根元の、細胞内に埋没した部分は鞭毛上部と構造が異なりこう呼ばれる。基底小体は周辺微小管が三連管(A小管、B小管、C小管)より成り、中心対微小管は存在しない。細胞分裂に関わる中心小体と同じ構造であり、実際に微小管形成中心(MTOC; microtubule organizing center)として機能するものもある。二個以上の基底小体を含む鞭毛装置では、その間を連結繊維と呼ばれる繊維構造が連結する場合が多い。
- 鞭毛根(flagellar root):基底小体に付随する微小管や繊維の総称。細胞内における鞭毛の位置を固定する役割があると思われる。
- 微小管性鞭毛根:微小管で構成された鞭毛根。鞭毛装置から伸びた微小管性鞭毛根は、細胞全体に広がって骨格微小管を兼ねる場合や、捕食装置の一部として機能する場合もある。特に高密度に微小管が配列したものは結晶性鞭毛根と呼ばれる。
- 繊維性鞭毛根:微小管よりも細い繊維系で構成された鞭毛根。電子顕微鏡では電子密度の高い領域として観察される。不等毛植物やプラシノ藻類では、リゾプラストと呼ばれる明瞭な縞模様を持ったものが観察される。
-
 (1)
(1) -
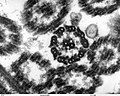 (2)
(2) -
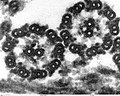 (3)
(3) -
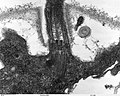 (4)
(4) -
 (5)
(5) -
 (6)
(6) -
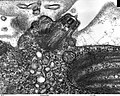 (7)
(7) -
 (8)
(8)








- (1) - (3):緑藻クラミドモナス(Chlamydomanas reinhardtii)の鞭毛断面
- (4) - (8):C. reinhardtii の鞭毛装置断面
- いずれも透過型電子顕微鏡(TEM)像
動作の種類
[編集]鞭毛の動作は大きく二種類の運動、鞭毛運動と繊毛運動とに分かれる。基本的に後者が可能な生物は前者も可能であり、運動の切り替えはカルシウムイオン濃度により制御されている。渦鞭毛藻やユーグレナ植物はいずれにも属さない独特の運動を行うが、その動作原理は未だ明らかでない。
- 鞭毛運動
- 繊毛運動
- 推進力を生み出す有効打(effective stroke)と、次の有効打の準備として鞭毛を元の位置に戻す回復打(recovery stroke)とを繰り返す運動。こちらは平泳ぎに近い。
生物の系統と鞭毛の修飾構造
[編集]従来、真核生物のは鞭型と羽根型(両羽/片羽)の2型に分類されていた。しかし「羽」と呼ばれて一括りにされてきた修飾構造には、分類群毎の差異がある事が電子顕微鏡の普及と共に判明してきた。従って、鞭型鞭毛や羽根型鞭毛という表現は、単に鞭毛の形状を表した便宜的なものにすぎない。
明瞭な修飾構造を持たない分類群
オピストコンタ(後生動物、菌類)、アメーボゾア、ケルコゾア、緑藻類など。
修飾構造を持つ分類群
- 褐藻類や珪藻類といった独立栄養の不等毛植物から、卵菌類やラビリンチュラのような従属栄養の生物までを含む。最も修飾を発達させたグループで、鞭毛の表面に管状マスチゴネマと呼ばれる小毛を持つ。この小毛は中空であり、基部・軸部・先端毛という三部構成となっている。管状小毛を持つ鞭毛は、鞭毛運動時に細胞に逆向きの推進力(鞭毛を前に向けて泳ぐ力)をもたらす。「ストラメノパイル」という名前はこの管状小毛に由来する(stramenopiles, straw;麦わら+pilus;毛)。
- 鞭毛は数種類の鱗片に覆われる。小毛を持つように見える種もあるが、これは毛状鱗片である。プラシノ藻の多くは鞭毛のみならず細胞本体の表面にも鱗片を持ち、それらの形態や組み合わせは属や種の同定に有用な形態形質である。
- 二本の亜等長 - 不等長鞭毛を持つ。長鞭毛が両羽、短鞭毛が片羽という組み合わせが多いが、これは属により様々である。長鞭毛の小毛は中空で、軸部と先端毛との二部構成である。ストラメノパイル同様、推進力を逆転させる働きを持つと言われる。
- ハプト藻類
- パブロバ亜綱では長鞭毛に小毛と鱗片を持つ。プリムネシウム亜綱では二本の鞭毛は等長もしくは亜等長であり、修飾構造を持たない。
- 鞭毛は片羽型。小毛は中空ではなく、またストラメノパイルのものよりも柔軟で推進力を逆転させる機能は無い。羽として認識される小毛の他に、さらに細かい毛が鞭毛全体を覆っている。
- 渦鞭毛藻類
- 横鞭毛は片羽型で、やはり中空ではない。縦鞭毛に小毛を持つ種もある。
- 複数本の鞭毛を持つものから無鞭毛のものまで様々であるが、多くの属では鞭毛のうちの一本(後曳鞭毛;recurrent flagellum)が細胞表面に付着し、後方に伸びる。この後曳鞭毛と細胞との間に波動膜を持つ種もある。一方、超鞭毛虫類は鞭毛に修飾を持たない。
渦鞭毛藻と同じアルベオラータである繊毛虫が持つ繊毛は、機能的、構造的に真核生物鞭毛と同じものである。
- また、水中の真核生物の分類群の中で紅藻類だけは、生活環を通して一切の鞭毛を持たない。有性生殖時に生じる精子も不動精子である。陸上生物では接合菌、子嚢菌と担子菌も一切の鞭毛細胞を生じない。種子植物の大部分もそうである。これらは二次的に鞭毛細胞を生じなくなったものと考えられる。種子植物ではソテツ類とイチョウ類だけが精子に鞭毛を持ち、祖先が鞭毛細胞を持つものであったことをうかがわせる。

細菌鞭毛
[編集]大腸菌をはじめとするバクテリア表面にみられる。直径20ナノメートル、長さ約十マイクロメートルのねじれた繊維。暗視野顕微鏡などの光学顕微鏡で観察することができる。真核生物のものと異なるのは、チューブリンからなる微小管ではなく、細菌では代わりにフラジェリンというタンパク質が重合して伸びた繊維でできていることである。真核生物の鞭毛とは運動機序が異なりダイニンの利用は見られない。
それぞれの繊維の付け根には回転モーターがあり、細胞内外のイオンの透過に共役した電気化学的ポテンシャルを運動エネルギーに変換することで回転する。そのためこのモーターの回転にはATPは必要ない。消費されるのは水素イオン濃度差である。このモーターの機構は電子伝達系によって駆動するATPaseと共通する部分が多い。フラジェリンのらせん状の繊維がこのモーターで回転すると、こうした微小な世界ではレイノルズ数が小さく水の粘性が高くなっているため、いわば粘っこい水の中にコルク抜きをねじ込むような形になり、細胞は高速で前進する。
回転モーターを除く鞭毛部分はIII型分泌装置とほぼ同様である。III型分泌装置を持つ細菌は比較的狭いグループに限られることから、鞭毛がIII型分泌装置に進化したとする見方が一般的だが、その逆とする説もある。
鞭毛繊維部分の構築は先端から行われる。これはIII型分泌装置が細胞外にたんぱく質を放出する際と同様の機構で、中空になっている鞭毛繊維部分の中をフラジェリンが通って先端に輸送される。
細菌鞭毛は、真核生物鞭毛と区別するために慣用的に「べん毛」と書かれることもある。
古細菌鞭毛
[編集]
広範囲の古細菌に存在する。繊維部分は細菌よりもやや細い直径10-15 nm、全長10-15 µmのねじれたタンパク集合体である。これも細菌と同様の機能を持ち、回転により移動力を得る。顕微鏡下では細菌鞭毛と殆ど見分けがつかず、見た目の違いは若干細い事、大抵は複数の鞭毛が束になっていることぐらいである。このため、1990年代中ごろまでは細菌鞭毛と同一の構造とみなされていた。しかしながら、鞭毛を構成するタンパク質に共通点は一切なく、両者は異なる起源を持つと考えられる。
古細菌の鞭毛を構成するたんぱく質は、古細菌自身やグラム陰性細菌が持つIV型線毛と類似が見られ、同様に根元から構築される。IV型線毛は付着のための器官で回転力は一切与えないが、これに回転モーターなどが追加され鞭毛を成している。駆動トルクはATPの加水分解により得ているが、エネルギー変換効率は水素イオンやナトリウムイオン濃度差をエネルギー源に利用する細菌に比べて著しく低く、6-10%程度と見積もられている。
Halobacterium salinarumにおいて詳細に観察されている。[1]
脚注
[編集]- ^ Yoshiaki Kinosita, Nariya Uchida, Daisuke Nakane, Takayuki Nishizaka(2016). “Direct observation of rotation and steps of the archaellum in the swimming halophilic archaeon Halobacterium salinarum.” Nature Microbiology 2016 Aug 26;1:16148
参考文献
[編集]- バイオダイバーシティ・シリーズ(3)藻類の多様性と系統:千原光雄 編 裳華房(1999)6章:鞭毛と鞭毛装置に見る多様性(井上 勲)
関連項目
[編集]外部リンク
[編集]この項目は、生物学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:生命科学/Portal:生物学)。 |