Lipnicovité (Poaceae) je čeleďjednoděložných rostlin z řádulipnicotvaré (Poales). Pro čeleď je povolen alternativní název Gramineae.[1] V češtině jsou obecně označovány jako trávy. Kromě typických trav bylinného vzrůstu sem však patří také dřevnaté bambusy, které mohou dorůstat až několika desítek metrů. Lipnicovité jsou charakterizovány hlavně kolénkatými stébly, štíhlými listy, jejichž báze obepíná stéblo jako pochva, a hlavně jedinečným typem květenství, jehož základem jsou tzv. klásky. Klásky jsou jedno- nebo vícekvěté jednotky podepřené dvěma listeny (plevami), každý z nenápadných kvítků v klásku je podepřen ještě pluchou a pluškou. Trávy jsou větrosprašné a produkují velké množství pylu. Ten může u citlivých lidí způsobovat alergické reakce („sennou rýmu“). Aby byl pyl snáze zachycen, je dvouramenná blizna trav dosti členitá (zpeřená). Plodem lipnicovitých je obilka, jejíž oplodí srůstá s osemením. V obilkách je dobře vyvinuté embryo a zásobní škrobnatý endosperm. U mnoha linií teplomilných trav došlo k přechodu k C4 typu fotosyntézy ze standardní fotosyntézy C3.[2][3][4]
Lipnicovité jsou velmi početná čeleď (přes 10 000 druhů v asi 700 rodech), ale je to také čeleď velmi významná ekologicky i ekonomicky. Travnaté louky a pastviny jsou základem pastevectví i zdrojem krmiva pro chov ustájených zvířat. Především však mezi trávy patří obilniny (globálně nejvýznamnější jsou kukuřice, pšenice a rýže), jejichž vařené, drcené, mleté a jinak zpracované obilky jsou nejvýznamnější zdroj lidské potravy. Využívají se však i k přípravě alkoholických nápojů – např. ječmen na výrobu piva. Obilniny patří mezi nejstarší domestikanty mezi rostlinami, přičemž sehrály významnou roli při přechodu lidstva k zemědělství. Další významná plodina z této čeledi je cukrová třtina, globálně nejvýznamnější cukrodárná rostlina. Lipnicovité mají i technický význam. Všestranně využitelným materiálem je sláma: používá se pro energetické účely, k výrobě papíru, jako podestýlka, ale jako ekologický a dobře izolující materiál i ve stavebnictví. Jako levný, ale pevný a lehký stavební materiál se používají i stébla bambusů. V podobě trávníků a druhů pěstovaných jako okrasné rostliny mají mnohé lipnicovité i rekreační a dekorativní roli. Na druhou stranu najdeme mezi travami i významné plevelné nebo agresivní invazní druhy poškozující původní ekosystémy.[2]
Trávy najdeme na všech kontinentech světa včetně Antarktidy. Obývají nejrůznější prostředí od tropických pralesů po pouště či tundry, řadě ekosystémů jako jsou savany, stepi, vysokohorské trávníky aj. dominují a určují jejich dynamiku. Nejstarší pozůstatky lipnicovitých rostlin známe z křídy. První trávy pravděpodobně rostly v podrostu gondwanských tropických pralesů. Pralesní prostředí je dodnes domovem trav z několika bazálních podčeledí. Tyto trávy mají často poměrně široké listy a někdy i nestandardně stavěná květenství a květy. V mírném pásmu (včetně území ČR) je z celkem 12 podčeledí trav zdaleka nejvýznamnější podčeleď lipnicové (Pooideae).[2][3]
Část stébla bojínku lučního: 1 – čepel; 2 – jazýček, 3 – listová pochva hořejšího listu, 4 – kolénko, 5 – stéblo nezakryté listovými pochvami, 6 – listová pochva (v horní části pootevřená)
Trávy mají zpravidla charakteristický „trávovitý“ vzhled, tj. tvoří často trsy štíhlých dlouhých listů, na dlouhých lodyhách (stéblech) jsou bohatá květenství nenápadných větrem opylovaných květů atd. Rostliny s tímto charakterem (pravé trávy, šáchorovité, sítinovité, orobincovité aj.) se v botanice a ekologii hromadně označují jako graminoidy,[5] v češtině se někdy používá neformální pojem „trávoidy“.[6][7]
Trávy jsou nejčastěji jednoleté, dvouleté či vytrvalé byliny, ale někdy to v důsledku intenzivní tvorby sklerenchymu kolem četných cévních svazků mohou být i netypické dřeviny, což platí zvláště pro bambusy, mezi kterými lze výjimečně nalézt i druhy chovající se jako opěrné liány.[4] Trávy dorůstají výšky od několika málo centimetrů až po více než 30 m u některých druhů bambusů.
Nadzemní stonky trav se nazývají stébla. Ta jsou často dutá nebo vyplněná dření (např. kukuřice nebo cukrové třtiny – třtinovce). U dutých stébel najdeme na příčném řezu pod povrchem jeden nebo dva kruhy cévních svazků doprovázených sklerenchymatickými pletivy zpevňujícími stéblo. V uzlinách stébel bývají nápadná kolénka, z nichž vyrůstají listy. V oblasti kolének je stéblo plné, uvnitř kolének je vytvořena spleť vzájemně propojených cévních svazků. Těsně nad kolénkem, krytý nejnižší částí listové pochvy, je prstenec dělivého pletiva, který spolu s intenzivně rostoucími vrcholovými partiemi pomáhá mladému stéblu růst do výšky a polehlému stéblu umožňuje znovu se zdvihnout tím, že se jeho buňky množí na straně orientované dolů. Stébla se většinou (mimo oblast květenství) nevětví, u bambusů a některých dalších trav ano. Kromě vyvýšených stébel s dlouhými články má většina trav i krátké přízemní až podzemní články schopné větvení, z nichž může regenerovat po pastvě, sečení nebo požárech. Vzácně jsou přízemní články ztloustlé v hlízky a fungují jako zásobní orgány (např. severoamerická strdivka Melica bulbosa).[3] Vytrvalé druhy mívají oddenky. Některé trávy jsou výrazně trsnaté, jiné rozvolněné, vytvářející dlouhé výběžky.[2]Klonálním rozrůstáním se do svého okolí mohou někteří jedinci během let kolonizovat rozsáhlé plochy a zřejmě se dožívat vysokého věku minimálně desítek, ale možná i stovek let.[3]
Několik příkladů různorodého habitu trav
Rákos obecný se do okolí rozrůstá oddenkovými výběžky
Listy vyrůstající z přízemních zkrácených článků dosti často tvoří přízemní růžice nápadné zvláště u trsnatých trav. Jsou jednoduché, střídavé, na stoncích skoro vždy dvouřadě uspořádané, většinou přisedlé, řidčeji (např. u většiny bambusů a u zástupců bazálníchpodčeledí) s tzv. nepravým řapíkem, tedy zúženou dolní částí čepele. Dolní část listu trav vytváří listovou pochvu obepínající stéblo, teprve výše odbočuje čepel. Čepele jsou celokrajné, nejčastěji úzké, čárkovité, někdy štětinovité či jehlovité, řidčeji až kopinaté nebo až vejčité. Žilnatina je převážně souběžná, zřídka dlanitá či zpeřená. Čepele mohou být ploché, někdy jsou žlábkovitého tvaru či svinuté, zřídka oblé. Na rozhraní pochvy a čepele je zpravidla přítomen jazýček různého tvaru či délky, který může být brvitý, chlupatý či lysý, roztřepený či celokrajný. Někdy může být redukovaný na věneček chlupů, např. u rákosu, nebo může být celkově zakrnělý až zcela chybět. Přízemní listy mohou výjimečně vytvářet drobné cibulky (např. lipnice cibulkatá, Poa bulbosa).[2]
Schéma vnitřní stavby listu kukuřice, C4 rostliny: kolem cévního svazku s xylémem (červeně) a floémem (oranžově) je vytvořena pochva, jejíž buňky (fialově) mají četné chloroplasty a v Calvinově cyklu produkují glukózu. Atmosférický CO2 je ale fixován v buňkách mezofylu (zeleně). List je kryt pokožkou (světle zelená), která je vespod vybavena průduchy (modře).
Pokožka listů je tvořena buňkami uspořádanými do řad ve směru podélné osy listu. Na listu se střídají řady větších (delších) a menších (kratších) buněk. Kratší buňky pokožky se často vyvíjejí v buňky obsahující fytolity, křemičitá tělíska složením odpovídající amorfnímu opálu (SiO2·nH2O). Tato tělíska snižují stravitelnost trav a chrání tak rostlinu proti herbivorům, zejména hmyzím, jejichž chitinovákusadla a další příústní orgány jsou tělísky rychle obrušovány. Efekt na mnohem odolnější sklovinuzubůobratlovců je spornější. Z krátkých pokožkových buněk mohou vznikat i jednobuněčné nebo dvoubuněčné trichomy trav. Průduchy trav jsou paracytické (kromě svěracích buněk jsou lemovány dalšími dvěma buňkami paralelními s buňkami svěracími).[3]
U trav došlo během evoluce nezávisle mnohokrát (více než 20×)[8] k přechodu z klasické C3fotosyntézy na fotosyntézu typu C4, která je energeticky náročnější, ale omezuje fotorespiraci, což zvyšuje efektivitu asimilaceCO2. V takovém případě se buňky listového mezofylu specializují na fixaci oxidu uhličitého pomocí fosfoenolpyruvátkarboxylázy do oxaloacetátu, který je redukován na kyselinu jablečnou nebo přeměněn na kyselinu asparagovou. Tyto kyseliny jsou následně transportovány do buněk tvořících pochvu kolem cévního svazku, kde je z nich CO2 opět uvolněn a využit enzymem RuBisCO v Calvinově cyklu. Žilnatina v listech C4 trav je v listu mnohem hustěji rozložená, aby kvůli efektivnímu a intenzivnímu transportu organických látek každá buňka mezofylu přímo sousedila s buňkou pochvy cévního svazku.[3] Většina C4 trav patří do podčeledi prosové. Z významných plodin jsou to kukuřice, proso, čirok nebo třtinovec, dále jsou to např. ježatka nebo rosička.[8] Mezi C4 rostliny patří zhruba polovina druhů trav a zároveň většina C4 rostlin patří právě mezi trávy.[9]
Kořeny trav bývají tenké, dlouhé a větvené, tvoří většinou hustou spleť. Primární kořen, který je první částí mladé rostlinky, která opouští obilku, bývá krátkověký (i když může vytrvávat i několik měsíců) a jeho funkci postupně přebírají další kořínky vyrůstající z děložního článku (mezokotylu) a později adventivní kořeny vyrůstající z oddenků a výběžků. Pro kořeny trav je typická schopnost generovat koncentrováním iontů v kořeni vysoký kořenový vztlak, hlavně v noci. Voda se pak dostává do xylému, stoupá vzhůru a rostlina je schopna tak znovu zaplnit roztokem cévy, v nichž došlo během dne k přerušení vodního sloupce (embolii). Kolem kořenů mnoha trav, obzvlášť xerofytů, se činností buněk kořene i bakterií asociovaných s kořenem vytváří vrstva stmelených půdních částic. U lipnicovitých včetně obilnin je častá arbuskulární endomykorhiza výrazně usnadňující příjem fosforu.[3]
Schéma klásku lipnicovitých. 1 – plevy, 2 – plucha, 3 – osina, 4 – pluška, 5 – plenky (2 okvětní lístky), 6 – tyčinky, 7 – svrchní semeník (zde srostlý ze 3 plodolistů, 2 jsou redukované, je přítomno 1 vajíčko), 8 – péřitá bliznová ramena.Dva rozkvetlé dvoukvěté klásky ovsíku vyvýšeného; jeden z kvítků v klásku ovsíku je oboupohlavný, druhý samčí: 1 – dolní pleva; 2 – stopka klásku; 3 – horní pleva; 4 – plucha horního kvítku (s krátkou, zde nepatrnou osinou); 5 – péřitá blizna; 6 – plušky; 7 – tyčinky; 8 – osina; 9 – nápadně osinatá plucha dolního kvítku; 10 – růžkovité plenky
Trávy jsou převážně rostliny s oboupohlavnými květy, někdy mohou být všechny nebo jen některé květy či klásky na rostlině jednopohlavné, případně se tvoří jednopohlavná květenství (např. kukuřice setá), pak jde o jednodomé rostliny.[2] Jen vzácně to jsou dvoudomé rostliny (např. některé americké lipnice).[4]
Pro trávy jsou velmi charakteristická složená květenství, jejichž základem je tzv. klásek, vícekvěté či redukcí jen jednokvěté (např. psineček, proso aj.) dílčí květenství, jehož některé květy však mohou být jalové (sterilní). Květy v klásku, je-li jich přítomno více, jsou uspořádány dvouřadě. Klásek je v typickém případě podepřen 2 plevami, což je vlastně zvláštní typ listenu, které jsou buď přibližně stejné, nebo různé. Plevy jsou uspořádány střídavě (i když blízko sebe a mohou se zdát vstřícné), proto rozlišujeme dolní a horní plevu. Pleva může být osinatá, či nikoliv. V některých případech je plev jiný počet, vzácně jsou zakrnělé nebo chybí (např. tajnička). Klásky většinou skládají další složená květenství, hlavně laty, hrozny, klasy, popř. stažené laty napodobující klas (lichoklas, např. bojínek).[2] U bambusů mohou z paždí dolních plev vyrůstat větévky nesoucí další klásky.[3]
Okvětí je u trav velmi redukované a nenápadné. Dva vnější okvětní lístky jsou srostlé a přeměněné v plušku, která je většinou zřetelně dvoukýlná a na konci dvouzubá.[2] Občas je však interpretována jako útvar listencovitého či dokonce smíšeného původu.[3] Někdy může být pluška redukovaná nebo chybí. Vnitřní okvětní lístky jsou přeměněny v plenky (lodiculae), které jsou nejčastěji 2, vzácněji 3, někdy chybí. Květ je podepřen pluchou, což je útvar listenového původu. Plucha může být někdy osinatá, řidčeji může být osin i více. Tyčinky, gyneceum a plenky jsou uzavřeny mezi pluchou a pluškou. V době rozkvétání plenky zduří, oddálí pluchu a plušku a umožní tyčinkám a bliznám vyklonit se z květů.[2]
Tyčinky jsou nejčastěji 3, méně často jich je 6, jejich počet může být dále redukován na 1–2 (ze 3) nebo 4 (ze 6), u některých zástupců z podčeledi Bambusoideae jich může být mnoho (několik desítek až 120).[4] Tyčinky jsou většinou volné, zřídka nitkami srostlé nebo srostlé s plenkami, jejich prašníky jsou vrtivé (snadno pohyblivé ve větru) a produkují relativně velké množství pylu. Opylení probíhá skoro vždy větrem (jde o anemogamní rostliny). Opylování hmyzem se předpokládá u bazální podčeledi Anomochlooideae.[3]Synkarpní gyneceum je srostlé ze 2, řidčeji 3 plodolistů, ale všechny plodolisty mimo jednoho se v průběhu vývoj květu redukují a výsledný pestík je již jednolitý útvar. Semeník je svrchní, obsahuje jediné vajíčko. Čnělka nese 2 či méně často 3 bliznová ramena, která jsou péřitá, aby lépe zachytávala pyl nesený větrem.[2][10]
Pylové zrno trávy v optickém mikroskopu; měřítko = 20 μm. Z Song et al. (2012)[11]
Zralá pylová zrna obsahují tři jádra (jedno vegetativní a dvě generativní). Jsou kulovitá nebo oválná a vybavená jediným pórem (jsou monoporátní). Kolem póru je zesílený prstenec. Ve stěně pylového zrna chybí jemné kanálky (strobikuly) přítomné u pylu většiny ostatních lipnicotvarých rostlin. Pyl trav je jinak morfologicky poměrně uniformní a jednotlivé podčeledi se na základě pylových zrn v podstatě nedají vzájemně rozlišit.[3] Pyl trav je jedním z nejvýznamnějších aeroalergenů (alergenů transportovaných vzduchem) a ve většině rozvinutých zemí světa patří k nejčastějším příčinám alergií, působí tzv. sennou rýmu. Mezi důvody mimořádné významnosti trav jako zdroje alergenů patří i to, že jsou hojné v mnoha městských a příměstských biotopech a během kvetení vypouštějí do větru velká množství pylu. Příznaky zahrnují rýmu a podráždění nosní sliznice, záněty spojivek, ale u citlivých osob až astmatické záchvaty. Odhaduje se, že příznaky senné rýmy sezonně trápí asi jednu pětinu populace Evropy a Severní Ameriky (z jiných oblastí většinou není dostatek spolehlivých údajů). Jako konkrétní makromolekuly vyvolávající alergické reakce se v pylu uplatňují hlavně profiliny (proteiny podporující polymeraci aktinu), polkalciny (proteiny vážící vápník), transferázylipidů a další. Velmi častá je zkřížená reakce na tyto enzymy pocházející z různých druhů trav, případně i z dalších rostlin.[12]
Plodem lipnicovitých je převážně obilka. Zřídka (u některých bambusů) zde může obilka přeměněná i na jiný typ plodu, např. může mít charakter bobule (se zesíleným a zdužnatělým oplodím) nebo oříšku (semeno je v oplodí uloženo víceméně volně).[4][13] Obilka může být okoralá, tj. za zralosti těsně obalená pluchou a pluškou, které s ní mohou srůstat (např. ječmen, oves), nebo neokoralá, nahá (např. žito, pšenice). Plucha, pluška, ale i plevy a jejich osiny, případně úlomky celého plodenství se podílejí na šíření semen (větrem, vodou, pomocí živočichů apod.). Osiny mohou svými hygroskopickými pohyby (tedy kroucením nebo narovnáváním v závislosti na vlhkosti) napomáhat i s průnikem obilek pod zem.[2]
Obilka je v podstatě derivát nažky, kde osemení srůstá s oplodím. Většina obilky je vyplněna zásobním pletivem (endospermem), jehož buňky během tvorby obilky akumulují škrob a proteiny a v době zralosti semene se rozpadají. Buňky povrchové, tzv. aleuronové vrstvy endospermu zůstávají životaschopné déle a v průběhu klíčení vylučují enzymy, které štěpí škrob endospermu na jednodušší sacharidy vstřebávané embryem.[3] Vedle endospermu je v obilce uloženo dosti vyvinuté embryo. Podrobná molekulárně-biologická analýza potvrzuje starší (ale spíše minoritní) názor, že jeho děloha je silně modifikovaná a vytváří dvě struktury, jednak tzv. koleoptile (též „coleoptile“), které kryje růstový vrchol (plumulu) se zárodky několika (2 u rýže a lipnice, 4 u kukuřice) prvních listů, a jednak tzv. štítek (scutellum).[14][3] Štítek je v přímém kontaktu s endospermem a zprostředkovává výživu klíčící rostlinky. Oblast mezi štítkem a koleoptile, které se během klíčení může prodlužovat, se nazývá mezokotyl. Naproti embryonálním listům a koleoptile je vytvořen kořínek chráněný koleorhizou. Při klíčení ze semene nejprve vyráží kořínek, koleoptile roste směrem vzhůru a chrání první listy, které záhy skrz něj prorostou. Koleoptile je proti listům bledší a po několika dnech odumírá.[2][3]
Trávy jsou velmi početnou čeledí a, jak je z předchozího patrné, v řadě znaků vykazují značnou variabilitu. Většina výše uvedených znaků dobře charakterizuje v mírném pásmu hojnou a důležitou podčeleď lipnicové a (často jako „výjimky“) jsou připomenuty variace, které lze najít u dalších početných nebo významných podčeledí jako jsou prosové, rýžové nebo bambusové trávy. Nejbazálnější podčeleď, Anomochlooideae, přitom zahrnuje rostliny, které netvoří klásky ani plevy (ačkoli listeny v květenství mají), nemají okvětí přeměněné v plenky nebo plušku, jejich listy mají relativně širokou čepel, některé z nich (Streptochaeta) mají 6 tyčinek a 3 blizny atd. V těchto ohledech také Anomochlooideae, a rod Streptochaeta zvlášť, patrně mnohem lépe ilustrují situaci u posledního společného prapředka všech trav nežli pokročilejší a evolučně úspěšnější skupiny. Mezi ancestrální (původní) znaky lipnicovitých, které se zachovaly napříč celou čeledí, tak patří znaky jako stébelnatý charakter nadzemních stonků, jediné vajíčko v jednopouzdrém pestíku, značně vyvinuté embryo se štítkem v obilce i obilka samotná jakožto typ plodu, stavba pokožky se dvěma střídajícími se typy buněk a s křemičitými tělísky nebo monoporátní pylová zrna s prstencem. Jiné ancestrální znaky (tropický vlhký stinný habitat, široké listy s nepravými řapíky, šest tyčinek, fotosyntéza typu C3 aj.) se u mnoha trav nezachovaly. Klásky sice mezi původní znaky trav nepatří, ale v evoluci se objevily dosti záhy – po odštěpení podčeledi Anomochlooideae. Klásky (a také květy s plenkami, pluškou a pluchou) tedy charakterizují všechny trávy kromě Anomochlooideae.[3]
Počet chromosomů v diploidní sestavě kolísá od 2n = 4 (rody Zingeria nebo Colpodium) až 266 (některé lipnice). Výchozí počet chromosomů je v rámci čeledi vzhledem k situaci u bazálních podčeledí a sesterských skupin nižší (podle různých odhadů 2n = 10–36).[3] Rozsáhlá studie genomů několika zástupců navrhuje scénář, kde brzy během evoluce trav došlo k modifikaci výchozího stavu (2n = 10) celogenomovou duplikací (2n = 20) a následnou reorganizací genomu na sestavu 2n = 24, která je u některých zástupců zachována dodnes, u jiných byla dále pozměněna.[15] U většiny trav z podčeledi lipnicové se v jádře nachází 2n = 14 velkých chromosomů (počty chromosomů se ale mohou lišit i mezi druhy v rámci rodu). U trav je častá polyploidizace, většinou jako důsledek křížení (allopolyploidie). Např. u pšenic stojí hybridizace spojená s polyploidií i za vznikem kulturních odrůd – nejčastěji pěstované pšenice jsou hexaploidní, mají celkem 42 chromosomů (přičemž 42 = 3 × 14). Velikost genomů trav je také velmi variabilní – od diploidního genomu o velikost 205 Mbp u rodu Oropetium po (opět diploidní) genom žita o velikosti 6,79 Gbp.[3] Velikost genomu hexaploidních pšenic je asi 16 Gbp.[16] Pro srovnání, genom rýže je relativně malý (400 Mbp).[15] Variabilita ve velikosti genomu se týká zejména nekódujících sekvencí. Z velké části jsou za ni zodpovědné retrotransposony, které se přednostně vkládají do kopií sebe sama a tvoří tak charakteristické úseky DNA.[3]
Trávy jsou rozšířeny po celém světě (včetně nejsevernějších oblastí Antarktidy a přilehlých ostrovů). Centrum rozšíření většiny podčeledí (bambusy, rýžové nebo prosové rostliny) je v teplých oblastech, bazální podčeleď Anomochlooideae je z tropické Jižní Ameriky. Lipnicové trávy jsou hojně rozšířené především v oblastech s mírným klimatem včetně Evropy a Severní Ameriky, což jsou oblasti s dlouhou botanickou tradicí. I proto jsou právě zástupci této podčeledi vnímáni jako „typičtí“ představitelé lipnicovitých. Některé druhy zasahují i do Arktidy a Antarktického pásma.[2]
Trávy se vyskytují v nejrůznějších ekosystémech od tropických pralesů po pouště. Dominují různým travnatým ekosystémům – tropickým savanám a stepím mírného pásu (pampy, prérie, eurasijské stepi). Jde o ekosystémy, které alespoň po část roku trpí nedostatkem srážek a přecházejí v polopouště až pouště. Mnohé trávy mají xerofytní adaptace a je mezi nimi mnoho druhů výrazně suchomilných, je zde ale i mnoho druhů mezofilních, druhů lesů mírného pásma i tropů, ovšem na druhou stranu i hodně bahenních rostlin a rostlin vodních kořenících ve dně (zblochany, rýže). Pobřežní rákosiny mohou být příkladem dalšího, často dosti rozsáhlého ekosystému, jemuž dominuje zástupce trav, v tomto případě rákos obecný. Vysokohorské (alpínské) trávníky jsou dalším typem extrémního prostředí obývaného primárně trávami. V takzvaných vyfoukávaných alpínských trávnících nacházíme v důsledku silného větru i sklonu terénu jen mělké, kamenité, neúrodné a vysýchavé půdy v zimě chráněné jen tenkou sněhovou pokrývkou a tedy vystavené i silnému mrazu.[17] Umělé luční ekosystémy a trávníky běžné v ČR a dalších evropských zemích vzniklé většinou pastvou a dnes udržované spíše sečením v mnohém stepi připomínají a trávy v nich dominují též.[2] K ekosystémům dominovaným travami patří také rozsáhlé bambusové lesy Jižní Ameriky a Asie. Spolu s dalšími rostlinami, například šáchorovitými, mají lipnicovité podstatný podíl i na vegetaci tundry. Odhaduje se, že 30–40 % souše je pokryto ekosystémy s převahou trav. Zdá se, že důležitým faktorem udržujícím trvalé bezlesí zejména v savaně jsou stepní požáry, k nimž trávy akumulací suché biomasy výrazně přispívají. Herbivoři (jako třeba pakoně v Serengeti) mohou spasením velkého množství trávy způsobit snížení intenzity požárů a následné rozšíření stromů na úkor travnaté savany. Travnaté ekosystémy jsou často druhově velmi bohaté, ale stupeň jejich ochrany je obecně dost nízký, ačkoli se jejich plocha lidskou činností významně snižuje. Bývají totiž vhodné pro rozšiřování pastevních a polních ploch.[3]
Obrovská rozmanitost ekosystémů obývaných travami a nutnost přizpůsobení se podmínkám, které v nich panují, se odráží na fyziologii, anatomii i morfologii lipnicovitých. Asi nejnápadněji je tato variabilita zřetelná na jejich listech. Xerofytní trávy jako např. kostřava ovčí, kavyly a řada dalších mají často vyvinutou schopnost svinutí listů do ruličky, přičemž směrem ven je orientována spodní strana listu se silnější kutikulou a lépe chráněná sklerenchymatickým pletivem. Uvnitř svinutého listu je pak chráněná svrchní strana listu. Průduchy se nacházejí právě zde, a to většinou v rýhách, které jsou na svrchním povrchu vyvinuty. Proudění vzduchu a tím i zvýšený výpar vody jsou dále omezovány trichomy. V buněčných stěnách buněk pochev cévních svazků je zvýšená koncentrace ligninu a suberinu, které dále snižují ztráty vody. Na regulaci svinutí listů se podílejí veliké specializované buňky epidermis, tzv. buliformní buňky. K dalším xerofytním adaptacím patří i tvorba kompaktních trsů, kde se ve spleti stébel a listů udržuje vlhčí vzduch, nebo rozsáhlý a hluboký kořenový systém. Naopak v listech vodních trav, např. zblochanů, najdeme rozsáhlé podélné aerenchymatické dutiny vyplněné hvězdovitými buňkami.[18]
Obzvlášť zástupci podčeledi lipnicové, ale i další trávy rostoucí v mírném a chladném pásu jsou schopny dobře se vyrovnávat s chladem a mrazem. Vzhledem k tomu, že led má větší objem než kapalná voda, způsobuje zmrznutí vody v buňkách jejich roztrhání a odumření. Během aklimatizace na nízké teploty mění odolné druhy složení membrán, protože ty v nízkých teplotách snižují svou fluiditu („tuhnou“), dále snižují kapacitu fotosyntézy, neboť v chladu dochází ke zpomalení biochemických reakcí fotosyntézy, ale ne ke snížení tempa excitacechlorofylu světlem, což vede k oxidačnímu stresu, a konečně také zvyšují v buňkách koncentraci osmoticky aktivních látek, což napomáhá zadržet v buňkách dostatek vody, které se může nedostávat kvůli pomalejšímu příjmu kořeny i zvýšené propustnosti membrán. Přežití krátkodobých (např. nočních) lehčích mrazů řeší mnohé lipnicovité rostliny uchováním vody v nestabilním přechlazeném stavu – v nepřítomnosti krystalizačních jader může voda zůstat tekutá i pod bodem mrazu. Navíc trsnatost přispívá k tepelné izolaci částí rostliny. Přežití dlouhodobých mrazů vyžaduje speciální adaptace, jako je přítomnost proteinů inhibujících vznik krystalů ledu v buňkách nebo naopak proteinů sloužících jako krystalizační jádra v mezibuněčných prostorech a na povrchu rostlin. Důležitým kryoprotektivním mechanismem je syntéza fruktanů, které stabilizují membrány, zlepšují osmotický stav buňky a snižují míru oxidativního stresu neutralizací reaktivních forem kyslíku.[19] Zcela extrémní prostředí obývá v severní Antarktidě a v Jižní Americe rostoucí metlice antarktická, která se nevyrovnává jen s velmi chladným podnebím, ale i se střídavým suchem a podmáčením, vysokou salinitou nebo vyšší intenzitou UV záření.[20]
Složkou potravy dinosaurů rodu Equijubus byly možná i trávy
Fosilní doklady evoluce trav jsou v zásadě trojího typu: kromě relativně vzácných makrofosilií (větších, okem viditelných zbytků) jsou to mikroskopická pylová zrna a křemičité fytolity. Většina makrofosilií (hlavně obilek a částí květenství) pochází z miocénu, kdy se trávy začaly intenzivně šířit, a z pozdějších období. Přesto jsou známé i doklady klásků a listů z předchozích období včetně raného eocénu (asi 55 miliónů let před současností). Ještě starší nálezy (např. asi 100 miliónů let staré pozůstatky listů a klásků v barmskémjantaru)[21] byly také interpretovány jako pozůstatky lipnicovitých, ale tyto závěry jsou některými odborníky zpochybňovány.[3] Další podobně staré fosilie z Myanmaru ale zahrnují i zkamenělá sklerociaparazitickéhouby připomínající dnešní paličkovici parazitující v kláscích.[22][23]
Pyl mnoha dalších jednoděložných rostlin je pylu trav podobný, proto jsou i tyto doklady bez náležité analýzy pomocí transmisní elektronové mikroskopie sporné. Tak např. příslušnost pylu nalezeného v křídových vrstvách k travám je nejistá, u jihoamerického a afrického pylu starého 57 miliónů let je již pravděpodobná. Nálezy pylových zrn trav vzhledem k značné uniformitě příliš neinformují o evoluci jednotlivých podskupin. Křemičité fytolity jsou nalézány relativně hojně. Z pozdní křídy Indie byly popsány části epidermis trav s fytolity v koprolitechtitanosaurníchsauropodů.[23][24][3] Kdy přesně se během křídy trávy vyvinuly, je ale značně nejisté, různé interpretace fosilií a různé odhady založené na genetických rozdílech mezi současnými zástupci poskytují variabilní výsledky od nejstarší křídy až po křídu mladší.[3] Křídové fytolity (které již lze přiřadit jednotlivým podčeledím) nicméně naznačují diverzifikaci čeledi během křídy a její starší původ.[24] Tento závěr podporují i výše zmíněné makrofosilní zbytky z Myanmaru, ale také v roce 2018 publikované nálezy epidermis trav s fytolity objevené v asociaci s ústní dutinou ornitopodního (hadrosauroidního) dinosaura Equijubus normani z období spodní křídy (asi před 113 až 101 miliony let).[25]
Geografický původ trav je vzhledem k současnému rozšíření bazálních linií i rozšíření nejstaršího fosilizovaného pylu trav hledán v oblasti Gondwany.[26] Bazální podčeleď Anomochlooideae je jihoamerická, ostatní skupiny mají důležité archaické centrum diverzifikace v Africe. Trávy byly po dlouhou dobu (až do miocénu) spíše rostliny lesního a pralesního podrostu. Jeden ze scénářů předpokládá kolonizaci Eurasie travami v období raného eocénu, možná prostřednictvím Indického subkontinentu, který býval součástí Gondwany, ale připojil se k Eurasijské desce. Podíl lesních druhů trav byl během eocénu stále vysoký. Později, až během oligocénu, byla kolonizována Severní Amerika.[26]
Travnaté ekosystémy se staly výrazně dominantními během miocénu (na různých kontinentech v různou dobu). Za nejvýznamnější faktory tohoto přechodu od lesnatých k travnatým ekosystémům jsou považovány klimatické efekty jako je aridizace klimatu a výraznější sezonalita a následná změna dynamiky požárů, které dále přispěly k šíření savan a stepí. Paralelní mnohočetný přechod k fotosyntéze typu C4 nastal až poté, hlavně v období před 8–3 milióny let, tedy relativně pozdě.[27][28] Příčiny tohoto jevu nejsou jasné, v tomto období nenastaly žádné výrazné globální změny v koncentraci CO2 nebo množství srážek (nízká koncentrace CO2 a sušší prostředí zvýhodňují C4 rostliny). Miocenní rozmach travnatých ekosystémů ovlivnil také koloběh křemíku, neboť ten je travami ukládán ve formě fytolitů v listech a dalších částech těla. Při tlení, požárech a především při pastvě je tento křemík uvolněn, mobilizován a zčásti transportován do jezer i moří, kde je využíván rozsivkami k tvorbě jejich schránek. Zvýšená sedimentace diatomitu v miocénu tak patrně souvisí s rozšířením travnatých ekosystémů (a velkých savčích herbivorů).[3]
Následuje přehled a popis jednotlivých podčeledí a podskupin řazených do čeledi lipnicovité.[3][9][29] (V českém názvosloví mají podčeledi rostlin koncovku -ové.)
Anomochloa marantoidea (Anomochlooideae)
Anomochlooideae: jsou to vytrvalé širokolisté trávy, listy jsou s nepravými řapíky. Nemají typické klásky. Tyčinek je 4 (Anomochloa) nebo 6 (Streptochaeta). Plenky chybí nebo jsou nahrazeny brvami. Čnělka je 1, blizny jsou 3 (Streptochaeta) nebo 1 (Anomochloa). Patří sem pouze Anomochloamarantoidea a tři druhy rodu Streptochaeta, všechny původem z tropů střední a jižní Ameriky.
(klad trav s klásky, angl. „spikelet clade“: všechny ostatní podčeledi tvoří monofyletickou skupinu vyznačující se klásky; v květech mají plenky)
Pharoideae: vytrvalé jednodomé trávy. Klásky skládají laty a jsou podepřeny 2 plevami, jsou jednokvěté a jednopohlavné, často v párech samčí se samičím nebo někdy jsou samičí samostatně. V klásku je plucha a pluška a 3 plenky (lodiculae). Tyčinek je 6 (samčí květy). Jsou známy 3 rody a asi 14 druhů, původem z tropických lesů většiny světa.
(klad trav se dvěma bliznami, angl. „bistigmatic clade“: všechny následující podčeledi mají dvě blizny čnící z pestíků; jejich klásky jsou zpravidla vícekvěté)
Puelioideae: vytrvalé trávy. Klásky jsou oboupohlavné a tvoří latu nebo hrozen a obsahují několik květů. Plevy jsou 2. Květy jsou jednopohlavné, samčí a samičí, tyčinek je 6, čnělky a blizny 2–3, plenky 3. Patří sem jen 2 rody a 11 druhů, všechny z tropických deštných lesů Afriky.
(klad BOP + PACMAD: všechny ostatní podčeledi patří do tohoto velkého kladu pojmenovaného akronymem z názvů jednotlivých podčeledí, za jejich společné znaky jsou považovány tři tyčinky, dvě plenky a listy bez nepravých řapíků, ale všechny tyto znaky mohou u některých skupin revertovat do původnější podoby, obzvláště u bambusů)
(klad BOP: zahrnuje následující 3 podčeledi; donedávna byl uváděn jako BEP, ale místo původního Ehrhartoideae se od r. 2016 používá označení Oryzoideae)[29]Dendrocalamus giganteus (Bambusoideae), největší z trav, na staré fotografii z 19. století; všimněte si lidské postavy vlevo od paty bambusu
Isachne nipponensis (Micrairoideae)rýžové(Oryzoideae): jsou to jednoleté nebo vytrvalé trávy, lesní i nelesní, popř. vodní. Listy jsou dvouřadě uspořádané, jazýček je membránovitý nebo je nahrazen lemem chlupů. Klásky skládají hrozen nebo latu a jsou oboupohlavné nebo jednopohlavné. Plevy jsou 2, někdy chybí. Často jsou v klásku 1–2 sterilní květy a fertilní jen 1, kde vzniká obilka. Tyčinky jsou většinou 3 nebo 6, vzácně 1, 2 nebo 4, čnělky a blizny jsou 2, plenky také 2. Nejvíc zástupců je v teplých oblastech, hlavně na jižní polokouli, ale vyskytují se i jinde. Je známo asi 17 rodů a 120 druhů, příkladem je rýže (Oryza) nebo tajnička (Leersia)
bambusové (Bambusoideae): jsou to vytrvalé, vzácně jednoleté byliny, které někdy dřevnatějí, typicky bambus. Klásky jsou jednopohlavné nebo oboupohlavné, skládají klasy, hrozny nebo laty. Plevy mohou chybět nebo je 1, 2 či více. Tyčinky jsou 2, 3 nebo 6, někdy mnoho, až 120. Blizny a čnělky většinou 2–3, zřídka 1. Plenky jsou většinou 3, zřídka chybí nebo někdy jich je 6 či mnoho. Je známo asi 84–101 rodů a asi 940–1320 druhů, rozšířeny jsou hlavně v tropech s přesahem do teplejších částí mírného pásu. V Evropě jako původní chybí.
lipnicové (Pooideae): jsou to jednoleté i vytrvalé trávy. Jazýček je většinou membránovitý. Klásky jsou většinou oboupohlavné, zřídka jednopohlavné a skládají klasy, hrozny nebo laty. Plevy jsou většinou 2, vzácně 1 nebo obě chybí, v klásku je 1 až několik fertilních květů. Pluška je většinou dobře vyvinuta, vzácněji je zakrnělá nebo chybí. Tyčinky jsou obvykle 3, vzácně 1 nebo 2, blizny většinou 2, vzácně 1 nebo 3, plenky jsou většinou 2, vzácně 3. Patří sem asi 3800 druhů v 177 rodech, jsou rozšířeny hlavně v mírném až boreálním pásmu, v tropech více jen v horách. Náleží sem většina trav rostoucích v ČR.
(klad PACMAD: zahrnuje zbývající podčeledi)
Aristidoideae: jednoleté nebo vytrvalé trávy, často trsnaté xerofyty. Jazýček je membránovitý nebo věneček chlupů. Klásky vytváří latu, jsou oboupohlavné, květy také, z nich fertilní, ze kterého se vytváří obilka, je v klásku jen 1. Plucha má 3 osiny. Tyčinky jsou 1–3, blizny a čnělky 2, plenky 2, řidčeji chybí. Patří sem 3 rody (Aristida, Sartidia, Stipagrostis) a asi 300–385 druhy, rozšířeny v teplejších částech světa, v Evropě jen málo.
prosové (Panicoideae): jednoleté nebo vytrvalé byliny, listy mají někdy nepravý řapík. Jazýček je membránovitý nebo nahrazen věnečkem brv. Klásky tvoří laty, klasy nebo hrozny, jsou oboupohlavné nebo jednopohlavné, rostliny pak mohou být jednodomé (někdy s oddělenými samčími a samičími květenstvími – kukuřice setá) nebo dvoudomé. Klásky často obsahují jeden fertilní květ, kde vzniká obilka, z druhého květu je sterilní plucha. Pluška je dobře vyvinutá nebo redukovaná až chybí. Tyčinky jsou 2 nebo 3, blizny a čnělky 2 (vzácně 1 nebo 3), plenky 2 nebo někdy chybí. Je známo asi 212 rodů a přes 3300 druhů, které jsou rozšířeny více v tropech a subtropech, o něco méně v mírném pásu. Příkladem je proso (Panicum) nebo kukuřice (Zea).
trsťové (Arundinoideae): většinou vytrvalé, vzácně jednoleté byliny, někdy dřevnatí. Jazýček je membránovitý nebo věneček chlupů. Klásky (i květy) jsou oboupohlavné, většinou skládají latu, vzácněji hrozen nebo klas. Fertilních květů, ve kterých vzniká obilka, je v klásku jeden až několik. Plevy jsou 2, osina je někdy osinatá, vzácněji mohou být osiny 3. Tyčinky jsou většinou 3, vzácně 1 nebo 2, čnělky a blizny jsou 2, plenky jsou také 2. Patří sem asi 14 rodů a 20–38 druhů, rozšířených od mírného pásu po tropy. Z trav vyskytujících se v ČR sem patří např. bezkolenec (Molinia) nebo rákos (Phragmites)
Micrairoideae: jejich jazýček je přítomen ve formě řady štětů. Klásky typicky dvoukvěté. Dvě nesrůstající čnělky. Známo asi 9 rodů a skoro 188 druhů s tropickým rozšířením.
trojzubcové (Danthonioideae): vytrvalé, řidčeji jednoleté trávy. Jazýček membránovitý nebo věneček chlupů. Klásky jsou oboupohlavné, tvoří latu, řidčeji hrozen nebo klas, jsou mnohokvěté a obsahují jeden nebo více fertilních květů, ze kterých vznikají obilky. Plevy na bázi klásku jsou 2. Pluchy jsou osinaté, osina je často trojlaná nebo jsou přítomny 3 osiny, pluška je dobře vyvinutá, někdy krátká. Tyčinky jsou 3, blizny a čnělky 2, plenky jsou 2. je známo asi 19 rodů a 270 druhů, které jsou rozšířeny různě po světě, více však v teplejších oblastech. V ČR to je např. rod trojzubec (Danthonia).
Chloridoideae: jsou to jednoleté nebo vytrvalé trávy, často suchomilné. Jazýček je membránovitý nebo věneček chlupů. Klásky skládají laty (či laty klasů), klasy nebo hrozny. Klásky jsou oboupohlavné nebo jednopohlavné (pak jsou rostliny jednodomé nebo dvoudomé). Plevy jsou 2, fertilních květů s obilkou je v klásku 1 nebo více. Tyčinky jsou 1–3, blizny a čnělky 2, plenky 2 nebo chybí. Je známo asi 1350 druhů, které jsou rozšířeny hlavně v tropech a subtropech, méně v mírném pásu. Příkladem je milička (Eragrostis).
Význam lipnicovitých rostlin pro lidstvo je nedocenitelný, neboť do této čeledi patří nejvýznamnější zemědělské plodiny, obilí. Uvádí se, že asi polovina energetického příjmu lidstva je právě ve formě obilnin.[31] Nejdůležitější obilniny jsou z globálního hlediska kukuřice, pšenice a rýže. Dohromady tvořila jejich celková globální sklizeň v roce 2019 (měřeno v tunách obilek) asi 90 % celkové sklizně obilí. V produkci vede kukuřice (asi 1,15 Gt v roce 2019), která je ale z podstatné části spotřebovávána na výrobu biopaliv (bioetanolu). Následuje pšenice a rýže (asi 765, resp. 755 Mt v roce 2019). Většina zbývající produkce připadá na ječmen (skoro 160 Mt v r. 2019). Na pátém a šestém místě je z hlediska globální produkce čirok (skoro 60 Mt) a různé druhy prosa (necelých 30 Mt). Následují oves (23 Mt), tritikale (14 Mt) a žito (13 Mt).[32] Další obilniny mají spíše lokální význam. Patří mezi ně např. slzovka obecná, rosička útlá nebo milička habešská (tef).[33]
Obilky jsou po odstranění pluch, plušek, případně plev nejčastěji buď vařeny vcelku (rýže, prosné jáhly, ječné kroupy) nebo mlety na mouku či drceny na šrot, krupici, vločky apod. a dále zpracovávány. Obilná (hlavně pšeničná, v menší míře žitná) mouka je základní surovinou pro přípravu tradičních druhů pečiva a těstovin. Pražené obilky (žito, ječmen), často ve směsi s kořenem čekanky, mohou posloužit i jako náhražka kávy (melta). Z obilek lze za různými účely získávat škrob (pšeničný, rýžový, kukuřičný aj.).[13]
Obilky ječmene jsou jednou z nejdůležitějších surovin pro výrobu piva. Během klíčení dojde k enzymatické přeměně škrobu na jednodušší sacharidy (např. maltózu), které jsou pak zkvašoványkvasinkami. Známé jsou i další alkoholické nápoje odvozené od obilovin, např. rýžové sake nebo režná vodka, což je žitný destilát. Z různých obilovin se připravuje také whisky.[2][13] Velký význam má produkce cukru (sacharózy) z cukrové třtiny. Té byly v roce 2019 celosvětově sklizeny skoro dvě gigatuny, kdežto cukrové řepy pěstované jako cukrodárná rostlina v ČR pouze něco málo přes 275 Mt.[32] Také cukr z cukrové třtiny může být zkvašován na alkoholické nápoje – z třtinového cukru je vyráběn pravý rum.[13]
Co se živočišné výroby týče, i zde hrají lipnicovité rostliny nezastupitelnou roli jako důležitá složka krmiv. V případě intenzivních chovů (nejčastěji prasata, drůbež) jsou zvířata běžně krmena granulovanými krmivy s podílem obilí (např. ve formě šrotu). Extenzivně chovaná zvířata (především hovězí dobytek, ale také koně, velbloudi, ovce nebo kozy) jsou pasena na (více či méně umělých) travnatých ekosystémech dominovaných různými pícninami; kromě trav se na jejich výživě podílejí i další rostliny (velký význam mají pro vysoký obsah proteinůbobovité pícniny).[13] Celková rozloha pastvin a luk je přitom obrovská: v roce 2015 činila skoro 5 miliard hektarů (asi 2/3 zemědělsky využívané plochy).[34] Zejména v oblastech, kde není pastva přes zimu možná, se píce suší ve formě sena nebo konzervuje jinými způsoby (senážování). Některé druhy a šlechtěné odrůdy pícninářsky významných trav mohou být na plochách určených ke zkrmování dosévány. Mezi nejvýznamnější pícniny z čeledi lipnicovitých patří v ČR luční zástupci rodů jílek, srha, bojínek, kostřava, ovsík, lipnice, psárka, psineček, trojštět a další. Jako pícniny se ale využívají i na polích pěstované obilniny, často ve směskách s bobovitými pícninami (např. jetel, vojtěška).[35] Význam v živočišné výrobě má i sláma ve formě podestýlky; je potom důležitou (provzdušňující) složkou hnoje.
Ukázky využití trav v potravinářství a zemědělství
Pečivo, mouka a další potravinářské výrobky z obilí
Sklizená cukrová třtina čeká na další zpracování
Naklíčený ječmen je zdrojem sladu, suroviny pro výrobu piva
Bambusové lešení v HongkonguDům se stěnami z balíků slámy v pokročilém stádiu výstavby
Ze slámy obilnin lze splétat nejrůznější předměty, jako jsou ošatky, rohože, klobouky nebo i dekorativní předměty. Využívá se také jako surovina pro výrobu papíru, kvalitní papír poskytuje např. sláma rýžová. Sláma nebo rákos nacházejí (spíše historicky) uplatnění též jako střešní krytina (došky), ale začátkem 21. století se sláma jako zajímavý stavební materiál vrací: lze ji využít dokonce při výstavbě levných, rychle sestavitelných, ekologických, dobře izolujících domů.[36][37] Rákos má vzhledem ke svým pevným dlouhým stéblům i další technická využití (např. rákosové rohože). Z pevných větévek květenství čiroku se mohou vyrábět tradiční „rýžové“ kartáče a košťata.[13] Rozmanité využití mají také dřevnatá stébla bambusů – jde o univerzální lehký, ale pevný technický materiál. Výhonky bambusů jsou důležitou součástí asijské kuchyně.[2][38] Biomasa vyprodukovaná travami může být různým způsobem využívána i jako energetická surovina – v nejjednodušším případě spálením k produkci tepla. Alkohol vznikající zkvašením cukrů z naklíčených obilek kukuřice a dalších obilnin nebo ze stébel třtiny lze využít i jako příměs benzínu do spalovacích motorů (bioetanol). Travní hmotu lze využít i v bioplynových stanicích.[39]
„Citronová tráva“, voňatka citronová, velmi intenzivně voní po citronech a využívá se k ochucení čajů, v kosmetice a parfumerii.[4] Široké použití v parfumerii, aromaterapii a léčitelství má také vetiverie ovsuchovitá, produkující esenciální olej známý jako vetiver.[40]
Kortadérie dvoudomá, dekorativní tráva s nápadným květenstvím
Travní směsi různého druhového složení (v ČR obsahující většinou jílek vytrvalý, kostřavu červenou, lipnici luční a psinečky)[4] se vysévají jako trávníky pro nejrůznější dekorativní a rekreační účely (např. v parcích a dalších plochách veřejné zeleně, na zahradách domků, na hřištích, podél silnic a cest). Celková rozloha trávníků není zanedbatelná, odhaduje se, že v USA se jejich plocha blíží 2 % rozlohy území tohoto státu. Využívají se druhy uniformního, pokud možno svěžího vzhledu dobře snášející sečení a různé formy stresu (sešlap, prosychání, zastínění).[41] Různé druhy trav včetně bambusů se pěstují jako okrasné rostliny. Příkladem mohou být některé drobnější kostřavy vytvářející kompaktní polštářkovité, někdy zajímavě sivě zbravené trsy, nebo naopak mohutné trávy s nápadným květenstvím jako je ozdobnice čínská, kortaderie dvoudomá, zmíněná vetiverie a další.[4] Sušená květenství mnohých trav se mohou stávat součástí suchých kytic. Dekorativní klásky či celá květenství má např. třeslice nebo chrastice kanárská (Phalaris canariensis).
Existují další, mnohdy obtížněji definovatelné a finančně vyčíslitelné způsoby, jimiž trávy přinášejí užitek. Travnaté ekosystémy (louky, stepi, savany) jsou často ekosystémy s vysokou biodiverzitou.[3] Mají se svou charakteristickou scenérií také svou estetickou hodnotu částečně se odrážející třeba v míře ekoturismu. Trávy také svými spletitými kořeny a oddenky obecně přispívají k ochraně půd před erozí a některé se využívají ke zpevňování písčin, svahů, rekultivaci obnažených půd apod.[42] V období „dust bowl“, katastrofálních prachových bouří na severoamerických Velkých planinách, byly různé trávy, mimo jiné rodů Bouteloa, Andropogon nebo i v Americe nepůvodní druhy žitňáků, masově využity při nápravě škod napáchaných příliš intenzivním zemědělstvím v této oblasti.[41]
Do čeledi lipnicovitých ale patří i řada významných plevelů a invazních rostlin. Z mnoha plevelů vyskytujících se v ČR uveďme např. úporný a oddenkovými výběžky se snadno rozrůstající pýr plazivý, chundelku metlici úzce spjatou s ozimými obilninami nebo ježatku kuří nohu, která je společně s dalšími druhy tohoto rodu a s béry jedním z celosvětově nejvýznamnějších plevelů vůbec. Ježatky i béry jsou totiž schopny zaplevelovat pole s nejrůznějšími plodinami v dost různorodých klimatických podmínkách (jsou to tzv. generalistické plevele).[43] Některé ježatky se uplatňují jako plevele i v rýžových polích.[44] V kulturní rýži roste plevelně i planá rýže Oryza rufipogon ilustrující další fenomén spojený s plevely: s rýží setou se může křížit a produkovat tak hybridní potomstvo, které může být obtížnějším plevelem než planý mateřský druh. Podobně se s čirokem obecným může křížit obtížný a místy invazní plevel čirok halabský (Sorghum halepense).[45] Zbavovat se plevelů blízce příbuzných pěstované plodině je obecně z řady důvodů obtížné („nenápadnost“ kvůli podobnému habitu, obdobný tvar a rozměr plodů a proto jejich ztížená separace, obdobná citlivost k herbicidům atd.). Další globálně významné plevele, které se stávají obtížnými spíše v teplých oblastech světa, jsou troskut prstnatý (Cynodon dactylon), kalužnice indická nebo Rottboellia cochinchinensis. Nejvýznamnějším způsobem zavlékání plevelů do nových oblastí světa je kontaminované osivo.[43] Invazní druhy trav zavlékané člověkem po celém světě někdy významně poškozují místní ekosystémy. Příklady mohou být eurasijský a severofrický sveřep střešní (Bromus tectorum) nebo ovsiřík štíhlý (Ventenata dubia), které jsou velmi agresivními invazními organismy v Severní Americe. Nejen, že konkurují místním druhům rostlin, ale jejich hromadící se suchá biomasa zde přispívá ke vzniku a šíření požárů.[46][47]
Planá pšenice jednozrnkaSamičí klas teosinte (nahoře), kulturní kukuřice (dole) a jejich hybrida (uprostřed)
Trávy jsou široce rozšířené rostliny tvořící semena s bohatými zásobami škrobu v endospermu. Obsahují i nezanedbatelné množství proteinů (např. lepku). Obilky lipnicovitých rostlin jsou tudíž cenným potravním zdrojem, který sehrál roli i v evoluci moderního člověka. Savanovými travinami s C4 metabolismem (namísto listů, plodů a jiných „C3“ zdrojů) se podle analýz obsahu izotopůuhlíku ve sklovině zubů začali předkové moderního člověka živit před 3,5–3,75 milióny let.[48][49]Domestikace obilnin pak byla důležitou součástí přechodu k zemědělství a podstatnou měrou tak napomohla odstartovat neolitickou revoluci.
Druhy, které se postupně začaly pěstovat (vesměs jde o jednoleté druhy, které investují velké množství energie do tvorby semen), byly lidmi sbírány a využívány již dlouho před počátkem zemědělství. Například zrnka divoké pšenice jsou doložena z archeologických nalezišť v Úrodném půlměsíci starých přes 19 000 let, ačkoli skutečné zemědělství se zde začalo rozvíjet nejspíše během relativně chladného a suchého mladšího dryasu před cca 12 000 lety.[50] Mezi důležité rané domestikanty patří v této oblasti diploidní pšenice jednozrnka (Triticummonococcum), tetraploidní pšenice dvouzrnka (T. turgidum subsp. dicoccum) nebo ječmen setý. Tyto trávy se zde začaly pěstovat před více než 10 000 lety a jejich pěstování se šířilo i do dalších oblastí. Zásadní událostí v evoluci kulturních pšenic byla hybridizace pěstované dvouzrnky a mnohoštětu Tauschova (Aegilopstauschii), která dala asi před 8 000 lety vznik nejvíce pěstovanému druhu, hexaploidní pšenici seté (T. aestivum subsp. aestivum).[51]
V případě kukuřice, která byla domestikována v oblasti jižního Mexika asi před 9 000 lety, došlo k výrazné změně morfologie samičích klasů: u divokého prapředka (tzv. teosinte) je klas dvouřadý s menším počtem (asi 10) velmi tvrdých obilek.[52][3] Domestikace rýže proběhla zhruba ve stejné době ve východní Asii. Když se její pěstování rozšířilo i do dalších oblastí (jižní Asie), docházelo zde ke křížení kulturní rýže s místními populacemi divoké rýže, což přispělo ke vzniku nových odrůd a forem. Žito a oves jsou obilniny, které sice byly domestikovány v oblasti Blízkého východu nedlouho po pšenici a ječmeni, ale díky své vyšší odolnosti vůči chladu nabyly významu až po rozšíření zemědělství do Evropy.[3]
↑PARK, Chris; ALLABY, Michael. A Dictionary of Environment and Conservation. Svazek 1. [s.l.]: Oxford University Press Dostupné online. DOI10.1093/acref/9780191826320.001.0001. (anglicky) DOI: 10.1093/acref/9780191826320.001.0001.
↑HROUDA, Lubomír. Trávy a jejich příbuzní napříč biotopy I. Systematika, fylogeneze, morfologie (úvod). Živa. 2010. Dostupné online.
↑HROUDA, Lubomír. Cena Živy III. Lubomír Hrouda: Trávy a jejich příbuzní napříč biotopy. iForum. 2011. Dostupné online [cit. 2021-07-10].
↑ abSAGE, Rowan F. A portrait of the C 4 photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, and Hall of Fame. Journal of Experimental Botany. 2016-07, roč. 67, čís. 14, s. 4039–4056. Dostupné online [cit. 2021-07-07]. ISSN0022-0957. DOI10.1093/jxb/erw156. (anglicky)
↑GARCÍA-MOZO, H. Poaceae pollen as the leading aeroallergen worldwide: A review. Allergy. 2017-12, roč. 72, čís. 12, s. 1849–1858. Dostupné online [cit. 2021-07-23]. DOI10.1111/all.13210. (anglicky)
↑ abSALSE, Jérôme; BOLOT, Stéphanie; THROUDE, Michaël. Identification and Characterization of Shared Duplications between Rice and Wheat Provide New Insight into Grass Genome Evolution. The Plant Cell. 2008-02-26, roč. 20, čís. 1, s. 11–24. Dostupné online [cit. 2021-07-09]. ISSN1532-298X. DOI10.1105/tpc.107.056309. PMID18178768. (anglicky)
↑KOČÍ, Martin. Katalog biotopů České republiky. Příprava vydání Chytrý, Milan; Kučera, Tomáš; Kočí, Martin. 1. vyd. Praha: Agebtura ochrany přírody a krajiny ČR, 2001. Kapitola Alpínské trávníky.
↑KRAEHMER, Hansjoerg; BAUR, Peter. Grasses: crops, competitors, and ornamentals. Příprava vydání Kraehmer, Hansjoerg. 1. vyd. Frankfurt am Main: Bayer AG, 2019. ISBN9781119416869. Kapitola Leaf.
↑SCHUBERT, Marian, et al. To Coldly Go Where No Grass has Gone Before: A Multidisciplinary Review of Cold Adaptation in Poaceae. Annual Plant Reviews. 2020, roč. 3, s. 523–562. Dostupné online.
↑AMOSOVA, Alexandra V.; BOLSHEVA, Nadezhda L.; ZOSHCHUK, Svyatoslav A. Comparative molecular cytogenetic characterization of seven Deschampsia (Poaceae) species. PLOS ONE. 2017-04-13, roč. 12, čís. 4, s. e0175760. Dostupné online [cit. 2021-07-25]. ISSN1932-6203. DOI10.1371/journal.pone.0175760. PMID28407010. (anglicky)
↑POINAR, JR., George O. Programinis burmitis gen. et sp. nov., and P. laminatus sp. nov., Early Cretaceous grass-like monocots in Burmese amber. Australian Systematic Botany. 2004, roč. 17, čís. 5, s. 497. Dostupné online [cit. 2021-07-09]. ISSN1030-1887. DOI10.1071/SB04002. (anglicky)
↑POINAR, George Jr.; ALDERMAN, Stephen; WUNDERLICH, Joerg. One hundred million year old ergot: psychotropic compounds in the Cretaceous?. Palaeodiversity. 2015, roč. 8, s. 13–19. Dostupné online.
↑ abPRASAD, V.; STRÖMBERG, C.A.E.; LEACHÉ, A.D. Late Cretaceous origin of the rice tribe provides evidence for early diversification in Poaceae. Nature Communications. 2011-09, roč. 2, čís. 1, s. 480. Dostupné online [cit. 2021-07-09]. ISSN2041-1723. DOI10.1038/ncomms1482. (anglicky)
↑WU, Yan; YOU, Hai-Lu; LI, Xiao-Qiang. Dinosaur-associated Poaceae epidermis and phytoliths from the Early Cretaceous of China. National Science Review. 2018-09-01, roč. 5, čís. 5, s. 721–727. Dostupné online [cit. 2021-07-09]. ISSN2095-5138. DOI10.1093/nsr/nwx145. (anglicky)
↑ abBOUCHENAK-KHELLADI, Yanis; VERBOOM, G. Anthony; SAVOLAINEN, Vincent. Biogeography of the grasses (Poaceae): a phylogenetic approach to reveal evolutionary history in geographical space and geological time: ANCESTRAL BIOGEOGRAPHY AND ECOLOGY OF GRASSES. Botanical Journal of the Linnean Society. 2010-04-19, roč. 162, čís. 4, s. 543–557. Dostupné online [cit. 2021-07-09]. DOI10.1111/j.1095-8339.2010.01041.x. (anglicky)
↑CERLING, Thure E.; HARRIS, John M.; MACFADDEN, Bruce J. Global vegetation change through the Miocene/Pliocene boundary. Nature. 1997-09, roč. 389, čís. 6647, s. 153–158. Dostupné online [cit. 2021-07-09]. ISSN0028-0836. DOI10.1038/38229. (anglicky)
↑STROMBERG, Caroline; MCINERNEY, Francesca. The Neogene transition from C3 to C4 grasslands in North America: Assemblage analysis of fossil phytoliths. Paleobiology. 2011, roč. 37. Dostupné online. DOI10.2307/23014716.
↑ abcdSORENG, Robert J.; PETERSON, Paul M.; ROMASCHENKO, Konstantin. A worldwide phylogenetic classification of the Poaceae (Gramineae) II: An update and a comparison of two 2015 classifications: Phylogenetic classification of the grasses II. Journal of Systematics and Evolution. 2017-07, roč. 55, čís. 4, s. 259–290. Dostupné online [cit. 2021-07-09]. DOI10.1111/jse.12262. (anglicky)
↑GRASS PHYLOGENY WORKING GROUP, kolektiv autorů. Phylogeny and Subfamilial Classification of the Grasses (Poaceae). Annals of the Missouri Botanical Garden. 2002, roč. 88. Dostupné online. DOI10.2307/3298585.
↑JOINT WHO/FAO EXPERT CONSULTATION, (kolektiv autorů). Diet, Nutrition an the Prevention of Chronic Diseases. Ženeva: WHO, 2003. 16 s. Dostupné online. ISBN92-4-120916-X.
↑ ab FAOSTAT. www.fao.org [online]. [cit. 2021-07-09]. Dostupné online.
↑CHEN, Ming-Hsu; KAUR, Prabhjot; DIEN, Bruce. Use of tropical maize for bioethanol production. World Journal of Microbiology and Biotechnology. 2013-08, roč. 29, čís. 8, s. 1509–1515. Dostupné online [cit. 2021-07-09]. ISSN0959-3993. DOI10.1007/s11274-013-1317-1. (anglicky)
↑ What Is Aromatherapy and How Does It Help Me?. Healthline [online]. 2018-05-15 [cit. 2021-07-24]. Dostupné online. (anglicky)
↑ abKRAEHMER, Hansjoerg. Grasses: crops, competitors, and ornamentals. Příprava vydání Kraehmer, Hansjoerg. 1. vyd. Frankfurt am Main: Bayer AG, 2019. ISBN9781119416869. Kapitola What makes grasses attractive ornamentals, and where?.
↑GIBSON, David. Grasses and Grassland Ecology. 1. vyd. New York: Oxford University Press Inc., 2009. ISBN978-0-19-852918-7. Kapitola 1.4 Grassland goods and services.
↑ abKRAEHMER, Hansjoerg; BELL, Carl. Grasses: crops, competitors, and ornamentals. Příprava vydání Kraehmer, Hansjoerg. 1. vyd. Frankfurt am Main: Bayer AG, 2019. ISBN9781119416869. Kapitola Dominance of Grasses as Weeds.
↑LEVIN, Naomi E.; HAILE-SELASSIE, Yohannes; FROST, Stephen R. Dietary change among hominins and cercopithecids in Ethiopia during the early Pliocene. Proceedings of the National Academy of Sciences. 2015-10-06, roč. 112, čís. 40, s. 12304–12309. Dostupné online [cit. 2021-07-09]. DOI10.1073/pnas.1424982112. PMID26371308.
↑CHARMET, Gilles. Wheat domestication: Lessons for the future. Comptes Rendus Biologies. 2011-03, roč. 334, čís. 3, s. 212–220. Dostupné online [cit. 2021-07-09]. DOI10.1016/j.crvi.2010.12.013. (anglicky)
↑ZOHARY, Daniel; HOPF, Maria; WEISS, Ehud. Domestication of plants in the Old World : the origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin. 4. vyd. Oxford: Oxford University Press 243 s. Dostupné online. ISBN978-0-19-954906-1, ISBN0-19-954906-0. OCLC761379401 Kapitola Cerals.
↑TIAN, F.; STEVENS, N. M.; BUCKLER, E. S. Tracking footprints of maize domestication and evidence for a massive selective sweep on chromosome 10. Proceedings of the National Academy of Sciences. 2009-06-16, roč. 106, čís. Supplement_1, s. 9979–9986. Dostupné online [cit. 2021-07-25]. ISSN0027-8424. DOI10.1073/pnas.0901122106. PMID19528660. (anglicky)

.jpg)

 Rákos obecný se do okolí rozrůstá oddenkovými výběžky
Rákos obecný se do okolí rozrůstá oddenkovými výběžky Kostřava sivá je trsnatá
Kostřava sivá je trsnatá Bambusy jsou dřeviny tvořící celé lesy
Bambusy jsou dřeviny tvořící celé lesy


 Jílek vytrvalý tvoří trsy dlouhých štíhlých listů
Jílek vytrvalý tvoří trsy dlouhých štíhlých listů Vodní Hygroriza aristata z Asie má neobvykle široké listy
Vodní Hygroriza aristata z Asie má neobvykle široké listy
.jpg)

.jpg)





 Kvetoucí bambus trnitý (Bambusa bambos)
Kvetoucí bambus trnitý (Bambusa bambos) Část květenství kukuřice seté, v němž jsou patrné četné dvoukvěté samčí klásky
Část květenství kukuřice seté, v němž jsou patrné četné dvoukvěté samčí klásky Klásky tomky vonné tvoří lichoklasy, jsou trojkvěté, ale jen jediný kvítek je plodný; má jen dvě tyčinky
Klásky tomky vonné tvoří lichoklasy, jsou trojkvěté, ale jen jediný kvítek je plodný; má jen dvě tyčinky Klásky ovsa hluchého mají nápadné plevy, zralé obilky jsou objaty pluškou a pluchou s nápadnou osinou
Klásky ovsa hluchého mají nápadné plevy, zralé obilky jsou objaty pluškou a pluchou s nápadnou osinou_Voss_(11497756443).jpg)



.png)

 Metlice Deschampsia antarctica na Petermannově ostrově (Antarktida)
Metlice Deschampsia antarctica na Petermannově ostrově (Antarktida) Kamýš písečný roste na suchých stanovištích včetně pobřežních písečných dun
Kamýš písečný roste na suchých stanovištích včetně pobřežních písečných dun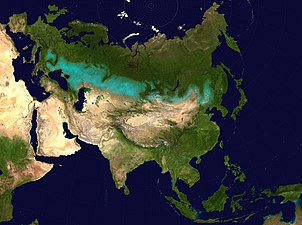 Pás eurasijských travnatých stepí (modravou barvou)
Pás eurasijských travnatých stepí (modravou barvou) Požáry pomáhají udržovat a šířit savany
Požáry pomáhají udržovat a šířit savany










 Pečivo, mouka a další potravinářské výrobky z obilí
Pečivo, mouka a další potravinářské výrobky z obilí Sklizená cukrová třtina čeká na další zpracování
Sklizená cukrová třtina čeká na další zpracování Naklíčený ječmen je zdrojem sladu, suroviny pro výrobu piva
Naklíčený ječmen je zdrojem sladu, suroviny pro výrobu piva Pasoucí se krávy
Pasoucí se krávy









 Obrázky, zvuky či videa k tématu lipnicovité na Wikimedia Commons
Obrázky, zvuky či videa k tématu lipnicovité na Wikimedia Commons