
Synapsiden
| Synapsiden Fossiel voorkomen: Laat-Carboon[1] – heden | |||||||||
|---|---|---|---|---|---|---|---|---|---|

| |||||||||
| Diverse uitgestorven en levende synapsiden | |||||||||
| Taxonomische indeling | |||||||||
| |||||||||
| Geen rang | |||||||||
| Synapsida Osborn, 1903 | |||||||||
| Afbeeldingen op | |||||||||
| Synapsiden op | |||||||||
| |||||||||
De synapsiden (Synapsida) zijn een groep gewervelde dieren die tot de Amniota behoren. De groep is genoemd naar een onderscheidend kenmerk: het bezit van slechts één onderste slaapvenster, een opening in de schedel achter elk oog. De vroege vormen van de groep worden wel informeel aangeduid met de term zoogdierachtige reptielen of zoogdierreptielen, hoewel ze geen reptielen zijn. Nog levende synapsiden zijn de zoogdieren.
Synapsiden waren de grootste gewervelde landdieren in het geologische tijdvak Perm, ongeveer 299 tot 251 miljoen jaar geleden. De groep onderging een enorme reductie tijdens de Perm-Trias-massa-extinctie. Tegen het einde van het Perm waren alle oudere vormen van synapsiden (de zogenaamde Pelycosauria) al verdwenen, zij hadden plaats gemaakt voor meer geavanceerde therapsiden. Na de Krijt-Paleogeen uitstervingsgolf radieerden de synapsiden opnieuw; ditmaal groeiden de zoogdieren uit tot de grootste land- en zeedieren.
Evolutie en afstamming
[bewerken | brontekst bewerken]Definitie
[bewerken | brontekst bewerken]
De term Synapsida werd in 1903 bedacht door Henry Fairfield Osborn. Osborn meende dat alle vroege amnioten in drie groepen te verdelen waren: de Anapsida zonder slaapvensters (fenestrae temporales), openingen in de zijwand van de achterste schedel; de Diapsida met twee slaapvensters en de Synapsida met slechts een onderste slaapvenster. De Synapsida fungeerden binnen deze opzet als een zogenaamde parafyletische groep, een "onderklasse" die een bepaalde groep afstammelingen niet omvatte, in dit geval de klasse zoogdieren.
- Schema schedelvormen volgens Osborn
-
 Schedel Anapsida (mogelijk de nog levende schildpadden)
Schedel Anapsida (mogelijk de nog levende schildpadden) -
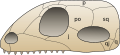 Schedel Synapsida (waaronder de huidige zoogdieren)
Schedel Synapsida (waaronder de huidige zoogdieren) -




In de moderne fylogenie worden geen parafyletische groepen gebruikt maar alleen monofyletische groepen, kladen die alle afstammelingen omvatten. De eerste exacte definitie van de Synapsida als klade werd in 1988 gegeven door Jacques Gauthier: de groep die de zoogdieren omvat en alle soorten nauwer verwant aan de zoogdieren dan aan de reptielen. Tot de klade Synapsida behoren dus per definitie de zoogdieren en de vroege, basale, synapsiden zijn per definitie geen reptielen. Hoewel de groep ernaar vernoemd is, is het bezit van slechts een onderste slaapvenster geen voorwaarde voor het behoren tot de Synapsida: alleen de afstamming is beslissend. In 2004 werd een wat moderner definitie gegeven door Robert Reisz die op soorten verankerd was. De Synapsida zijn dan de groep die Homo sapiens, de mens, omvat en alle soorten nauwer verwant aan Homo dan aan Iguana iguana, de groene leguaan. Sommige onderzoekers vinden het toepassen van de mens in dit soort definities wat pretentieus. In 2018 werd een meer "prehistorische" definitie gegeven: de groep die Dicynodon lacerticeps Owen 1845 omvat en alle soorten nauwer verwant aan Dicynodon dan aan Lacerta agilis Linnaeus 1756, de zandhagedis. De gebruikte soorten zijn slechts willekeurige vertegenwoordigers van hun groepen.
Ontwikkeling
[bewerken | brontekst bewerken]In het Carboon
[bewerken | brontekst bewerken]
In het Carboon, tijdens het Bashkirien, moeten de Synapsida zich al afgesplitst hebben. Dat weten we omdat toen al een soort van hun zustergroep de Sauropsida leefde, namelijk Hylonomus. De synapsiden moeten dus ook al bestaan hebben. Inderdaad is er een synapside vorm uit die tijd bekend: Protoclepsydrops. Daarvan zijn echter slechts beperkte resten gevonden zodat ook wel ontkend is dat het echt om een synapside gaat. De Jogginsformatie waarin beide vormen gevonden zijn, is vermoedelijk 318 miljoen jaar oud. Mogelijke voetafdrukken van synapsiden zijn nog een miljoen jaar ouder en zekere afdrukken dateren van 315 miljoen jaar geleden.

Later in het Carboon ontwikkelden zich soorten die met zekerheid als synapside te beschouwen zijn, op grond van complete gevonden skeletten, zoals Archaeothyris florensis en Echinerpeton intermedium. Zij hebben een ouderdom van 307 miljoen jaar. Uiterlijk leken deze dieren oppervlakkig op moderne hagedissen omdat ze ook gespreide poten hadden en een vrij kleine omvang. Ze leefden in de steenkoolwouden van Noord-Amerika. Binnen enkele miljoenen jaren waren de synapsiden de belangrijkste diergroep op Aarde en al tijdens het Carboon moet er zich een aanzienlijke diversificatie hebben voorgedaan met meerdere afstammingslijnen. Archaeothyris bijvoorbeeld is helemaal niet zo basaal maar staat vrij hoog in de stamboom zodat andere groepen Synapsida zich ook al ontwikkeld moeten hebben.
De Caseasauria kunnen zich al tijdens het Bashkierien hebben afgesplitst en een van hun takken waren de Caseidae, de eerste synapside planteneters. Hun zustergroep waren de Eupelycosauria. Tijdens het Moscovien moeten zich daarbinnen een reeks volgende groepen hebben afgesplitst, eerst wellicht de Varanopidae, dan de Ophiacodontidae en daarna de Edaphosauridae. De hoofdstam werd voortgezet door de Sphenacodontia waarvan de Sphenacodontidae een eerste afsplitsing waren. De oudste sphenacodontiden zijn gevonden in lagen van het Kasimovien. Hun zustergroep zijn de Therapsida die dus al rond 307 miljoen jaar geleden bestaan moeten hebben. Een probleem hierbij is dat dit een enorm lange verborgen afstammingslijn veronderstelt want de oudste zekere therapside fossielen duiken pas vijfendertig miljoen jaar later op. Er zijn verschillende verklaringen hiervoor geopperd.
Een eerste mogelijkheid is dat de vroegste therapsiden nog heel lang op basalere vormen bleven lijken zodat ze simpelweg niet als therapside herkend worden. Everett Olson verdedigde dit standpunt in een reeks artikelen tussen 1953 en 1992. Hij meende zelfs zulke soorten te kunnen aanwijzen. Dit verklaringsmodel wordt in toenemende mate onwaarschijnlijk. Verfijndere fylogenetische analyses maken het steeds duidelijker wat de subtiele kenmerken zouden moeten zijn die vroege therapsiden zouden bezitten. Olsens vermeende vroege therapsiden tonen die eigenschappen niet en worden tegenwoordig algemeen als kandidaat afgewezen. Een tweede verklaring is dat de Sphenacodontidae parafyletisch zijn en dus in feite niet één aftakking zouden vertegenwoordigen maar een reeks afsplitsingen waarvan dan de laatste pas de Therapsida waren. Zoiets veronderstelt een hoge mate van parallelle evolutie, het apart ontwikkelen van gelijkende kenmerken door verwante groepen. Het is in beginsel altijd mogelijk dat zoiets zich voordoet maar zonder nieuwe vondsten zullen de gangbare fylogenetische methoden dit niet kunnen ontdekken omdat die juist stambomen proberen te berekenen die de minste mate van parallelle evolutie veronderstellen. Een laatste mogelijkheid is dat therapsiden in een gebied geëvolueerd zijn waaruit men eenvoudigweg nooit fossielen gevonden heeft. Dat lijkt de beste verklaring te zijn want algemeen wordt aangenomen dat allopatrie, afsplitsing door geografische isolatie, het belangrijkste mechanisme is dat nieuwe taxa voortbrengt. Dit probleem staat in nauw verband met het vraagstuk wat de plotse overgang tussen een basale en een therapside fauna kan verklaren, later in het Perm.
In het Perm
[bewerken | brontekst bewerken]De basale fauna in het vroege Perm
[bewerken | brontekst bewerken]
Tijdens het Vroeg-Perm tot en met het Kungurien maakten Caseidae, Varanopidae, Ophiacodontidae, Edaphosauridae en Sphenacodontidae een bloeiperiode door. Hun fossielen zijn vooral gevonden in Noord-Amerika en Europa. Deze groepen ontwikkelden voor het eerst echt grote landdieren die zeker tot de Amniota behoren, hoewel gelijktijdig de Diadectomorpha ook een grote omvang bereikten en deze soms als amnioten worden beschouwd. Bekende grote soorten zijn de carnivoor Dimetrodon, de herbivoor Edaphosaurus en de vleeseter Varanosaurus. De sphenacodontide Dimetrodon — de enige basale synapside die bekend is bij een groter publiek — en Edaphosaurus waren in het bezit van een rugkam, die mogelijk een rol speelde bij regulering van de lichaamstemperatuur. Dit duidt erop dat de stofwisseling nog niet al te hoog lag. De ophiacodont Varanosaurus leek, zoals de naam al zegt, enigszins op een hedendaagse varaan. In het algemeen behielden de poten een spreidstand zoals bij huidige koudbloedige reptielen. Synapsiden met een dergelijke "ontwikkelingsgraad" worden traditioneel "Pelycosauria" genoemd. Het gaat hier echter niet om een natuurlijke groep daar de verschillende "pelycosauriërs" in meer of mindere mate verwant zijn met de meer afgeleide groep van de Therapsida.

Zo'n 270 miljoen jaar geleden, tijdens het Roadien, deed zich een grote omwenteling voor van de faunae. De Caseidae en de Varanopidae hielden het nog uit tot in het Capitanien maar andere basale groepen stierven volledig uit. Uit de periode daarna zijn in Rusland en Zuid-Afrika grote hoeveelheden therapside fossielen gevonden. Het is een zeer omstreden vraagstuk wat de aard is van deze verandering. De eerdere basale groepen zijn gevonden in een vrij nauwe band die van New Mexico via Texas door Canada naar West-Europa voert. Doordat de continenten toen verenigd werden in Pangea, lagen die gebieden indertijd direct aaneengesloten ter hoogte van de evenaar. De nieuwere vondsten liggen in een geografisch afgescheiden zones die zich tijdens het Perm veel noordelijker of zuidelijker op hogere breedtegraden bevonden, wat Rusland betreft ver op het Noordelijk Halfrond en Zuid-Afrika ver op het Zuidelijk Halfrond. Maar er bestaat niet alleen een geografische kloof maar ook een hiaat in de tijd. De oudste Russische vondsten zijn mogelijkerwijs twee tot drie miljoen jaar jonger dan de jongste Amerikaanse. Deze temporele kloof wordt wel Olson's Gap genoemd. Sommige onderzoekers ontkennen echter dat die kloof werkelijk bestaat en vermoeden foute dateringen; daarbij bestaat er een mogelijke oudere Amerikaanse therapside: Tetraceratops. De hypothese van Olson's Gap is verwant met de veronderstelling dat zich op het eind van het Kungurien ook een massa-uitsterven heeft voorgedaan, Olson's Extinction genaamd. Deze spectaculaire hypothese is op haar beurt ook weer ontkend; sommige gegevens zouden wijzen op een geleidelijke vervanging van de fauna. Doordat de vindplaatsen zover uit elkaar liggen, kan de abrupte overgang een illusie zijn en verder is het mogelijk dat de Therapsida zich juist op het Zuidelijk Halfrond ontwikkeld hebben en daar veel eerder voorkwamen.
De therapside fauna in het middelste en late Perm
[bewerken | brontekst bewerken]
Tijdens het Midden-Perm, het Guadalupien, deed zich een grote radiatie voor van de Therapsida. Als eerste groep splitsten zich de Biarmosuchia af. Daarop volgden de Dinocephalia en daarna de Anomodontia en de Gorgonopsia. De meest afgeleide tak, die zich al vroeg ontwikkelde, waren de Therocephalia. Hun zustergroep waren de Cynodontia die er dus ook al geweest moeten zijn, zij het weer in een verborgen ontwikkelingslijn.

De nieuwe fauna zag er heel anders uit dan de oude. De basale groepen moeten weinig actieve vormen geweest zijn met een lage soortendichtheid, relatief grote populaties en een trage evolutie. De therapsiden waren duidelijk veel actiever. Ze stonden meer geheven op hun achterpoten. Waarschijnlijk lag hun stofwisseling veel hoger, wellicht samengaand met de ontwikkeling van een vacht. Daardoor waren hun populaties kleiner en evolueerden ze sneller zodat er veel meer soorten waren. De vormenrijkdom was ook groter in de zin dat er meer morfologieën waren, typen lichaamsbouw. De Therapsida vulden de meeste niches voor grote dieren op. Naar huidige begrippen waren therapsiden in bouw ook heel bizar. Vooral de kopuitsteeksels waarmee kennelijk gevechten binnen de soort werden uitgevochten gaven de dieren een heel vervreemdende fysiognomie. Het grote publiek is tegenwoordig aardig op de hoogte van het uiterlijk van dinosauriërs maar heeft nog geen kennisgenomen van het zeker zo spectaculaire voorkomen van zijn nauwere verwanten uit het late Perm.

In Rusland leefden reusachtige carnivore soorten zoals de bijna vier meter lange gorgonopiër Inostrancevia, die in het bezit was van grote sabeltanden, en grote planteneters als de dinocefaal Estemmenosuchus. In Zuid-Afrika waren synapsiden zowel de voornaamste carnivoren als herbivoren. Tot de eerste groep behoorde onder andere Lycaenops, een kleine verwant van de Russische Inostrancevia. Sommige dinocefalen, de Brithopia, waren vleeseters. De reusachtige dinocefaal Moschops, die tot een ton zwaar kon worden, behoorde tot de plantenetende vormen, de Titanosuchia. De soortenrijkste groep planteneters waren de anomodontiërs en daarbinnen vooral de Dicynodontia. De Therocephalia vulden de niches van kleine vleeseters.
Het snelle evolutietempo uitte zich in een belangrijke faunawisseling op het eind van het Guadalupien: de Dinocephalia stierven uit en werden in de rol van grote landdieren vervangen door Neotherapsida.
De synapsiden deelden hun leefgebied in het Perm met grote amfibieën, en vroege reptielen behorend tot de Lepidosauromorpha en Archosauromorpha. De reptielen bleven echter klein en de Synapsida waren de dominante groep gewervelden, althans op het land: voor zover bekend waren er in deze fase geen echte zeedieren onder de Synapsida.
Aan het einde van Perm, 252 miljoen jaar geleden, deed zich een massaal uitsterven voor, het ernstigste uit de recentere wereldgeschiedenis: de Perm-Trias-massa-extinctie. Deze was vermoedelijk een gevolg van de vorming van de Siberische Trappen, gigantische vloedbasalten. Door deze langdurige vulkanische activiteit ontstond er een ernstig broeikaseffect dat de temperaturen op Aarde met 10 °C deed stijgen. Ongeveer 90% van alle diersoorten stierf uit. Juist de synapsiden werden niet gespaard omdat ze als grotere diersoorten hoger in de voedselketen stonden en dus als eerste het loodje legden als die ineenstortte. Vermoedelijk waren niet-therapside Synapsida op dat moment al uitgestorven maar mochten er nog restpopulaties van bestaan hebben, overleefden ze de extinctie in ieder geval niet. De Therapsida zelf kregen zware klappen. De meeste van hun afstammingslijnen eindigden. Enkele soorten daarvan bleven echter in leven en daaruit ontstonden de latere vormen.
Eigenaardig is dat de belangrijkste overlevende groep, de Cynodontia, tot die tijd een tamelijk onbelangrijke tak vormden. De oudste cynodonten zijn gevonden in het Wuchiapingien. Vermoedelijk ontstonden ze eerder en dat hun fossielen niet gevonden zijn, wijst erop dat ze maar een klein deel van hun fauna moeten hebben uitgemaakt.
In het Trias
[bewerken | brontekst bewerken]
Er waren twee afstammingslijnen die het uitsterven overleefden: de anamodontische Dicynodontia en de Cynodontia. De eerste groep omvatte in het Trias zwaargebouwde planteneters zoals Placerias, bekend van de BBC-serie Walking with Dinosaurs. Ze zijn genoemd naar een paar slagtanden in hun bovenkaken. De rest van het gebit was grotendeels vervangen door een hoornen snijrand. Ook Lystrosaurus behoorde hiertoe, de enige dicynodontiër die zowel uit het Perm als uit het Trias bekend is. Dit dier was direct na de extinctie zo wijdverspreid over Afrika, Zuid-Amerika, Rusland en zelfs Antarctica dat men wel van een globale "lystrosaurusfauna" spreekt, waarbij tot 90% van de gevonden fossielen van dit taxon is. Bij gebrek aan concurrentie kon het zich over de hele planeet verbreiden en alle latere dicynodontiërs, met uitzondering van Kombuisia, zijn nauw aan hem verwant. Ze werden echter niet meer zo soortenrijk als tijdens het Perm. De cynodonten waren kleiner en veelal vleeseters. Er waren basale vormen onder zoals Thrinaxodon. Een meer afgeleide deelgroep waren de Eucynodontia met als bekendste vroege soort Cynognathus.

In het Vroeg-Trias waren de synapsiden nog steeds de dominante dieren, hoewel ze in toenemende mate concurrentie ondervonden van de Archosauria. Het is omstreden wat hiervan de oorzaak was. Een mogelijkheid vormt dat de archosauriërs beter aangepast waren aan het droge klimaat dat tijdens het Trias heerste. Met hun schubbige huid en hun vermogen ureum droog af te scheiden verloren ze minder water. Een andere hypothese wil dat de archosauriërs de Synapsiden juist op hun sterkste punten aftroefden. Uiteindelijk ontwikkelden de archosauriërs een nog hogere stofwisseling, evolueerden een superieur ademhalingssysteem en gingen nog hoger op hun poten staan. Ze wisten ook meer niches te cultiveren. Archosaurische pterosauriërs kozen als eerste gewervelden het luchtruim. Crurotarsi raakten veel eerder dan synapsiden aangepast aan een leven in het water. Desalniettemin bleven Dicynodontia lange tijd de niches voor grote planteneters vullen. In 2008 waren er al 119 soorten van benoemd. De Synapsida verbeterden ook hun goede eigenschappen. Ze kregen een sneller metabolisme, nu met zekerheid ondersteund door een vacht, en gingen soms ook kaarsrecht op de poten staan. Daarbij werden ze, wat hun maximale omvang betreft, nog eens een orde van grootte zwaarder dan tijdens het Perm. De dicynodont Lisowicia die op het eind van het Trias leefde, had een gewicht van negen ton.

De Cynodontia begonnen ook een andere openstaande niche in te nemen: die van kleine warmbloedige gewervelde. Omdat kleine dieren snel afkoelen, waren daar zeer gespecialiseerde aanpassingen voor nodig. Ze ontwikkelden een dichte vacht en scherpe zintuigen gecombineerd met een gedifferentieerd gebit en een groot brein om energierijk voedsel te vinden en te verwerken. Zulke warmbloedige diertjes met snorharen en een goede reuk en gehoor konden ook 's-nachts actief zijn en zo de archosauriërs ontlopen. Al 246 miljoen jaar geleden was als zustergroep van de Cynognathia de eucynodonte klade Probainognathia ontstaan die zeer soortenrijk was en daarbinnen splitsten zich uiteindelijk de Mammaliamorpha af, de groep waartoe ook de zoogdieren behoren. Vroege mammaliamorfen waren de Tritylodontidae.
Op het eind van het Trias, 201 miljoen jaar geleden, was er weer een groot uitsterven, de Trias-Jura-extinctie. De Dicynodontia stierven toen wellicht geheel uit. De cynodonten bleven echter bestaan, hoewel veel deelgroepen verdwenen. Hun "miniaturisering" had het grote voordeel dat ze minder gevoelig werden voor massa-extincties. Groepen kleine dieren zijn vaak heel soortenrijk zodat de kans groter wordt dat er een het uitsterven overleeft. Hun populaties zijn ook groter, met hetzelfde effect. Er zijn altijd wel insecten die catastrofes zoals een meteorietinslag of vulkaanactiviteit overleven en die kunnen weer gegeten worden door kleine insecteneters. Het uitsterven is echter ook wel geleidelijker gezien als het gevolg van een concurrentiestrijd met de eerste dinosauriërs zoals Coelophysis. Binnen de Mammaliamorpha hadden zich ondertussen de Mammaliaformes ontwikkeld. Uit zulke muisvormige diertjes als Morganucodon zouden zich in de loop van miljoenen jaren de zoogdieren ontwikkelen — traditioneel werden de Morganucodontidae zelf als vroege zoogdieren gezien. Het is mogelijk dat zoogdieren in eigenlijke zin, afstammelingen van de laatste gemeenschappelijke voorouder van de moderne soorten, zich al in het Trias hadden afgesplitst maar hun fossielen zijn nooit gevonden.
Jura en Krijt
[bewerken | brontekst bewerken]Tijdens de Jura waren vermoedelijk bijna alle synapsiden vrij kleine dieren. De niches voor grotere dieren werden opgevuld door de dinosauriërs. Dat betekent echter niet dat ze een onbelangrijk deel van de fauna vormden. Er waren veel soorten, er ontwikkelden zich vele groepen zoogdieren met allerlei evolutionaire vernieuwingen en ze overtroffen de dinosauriërs bijna zeker in het aantal individuen.
In het begin van de Jura bleven basale Mammaliaformes de talrijkste synapsiden. Een belangrijke groep die in de middelste Jura opduikt, waren de Docodonta die complexe kiezen hadden. Zulke tanden vormen overigens de belangrijkste informatiebron over de synapside evolutie tijdens deze periode want hele skeletten zijn uiterste schaars. Een zeldzame uitzondering is het fossiel van de grote docodont Castorocauda.
Uit het vroege Krijt zijn er aanwijzingen dat er toch nog Dicynodontia overleefden in de vorm van stukken schedel met slagtanden die in Australië gevonden zijn. Op het eind van die periode werden zoogdieren gemiddeld wat groter. De catastrofe aan het eind van het Krijt, wellicht veroorzaakt door de inslag van een grote meteoriet, roeide ook veel synapsiden uit. Verschillende afstammingslijnen moeten het massauitsterven echter doorstaan hebben.
Cenozoïcum
[bewerken | brontekst bewerken]De ramp roeide echter bijna alle dinosauriërs uit, met uitzondering van de vogels. Dit gaf de synapsiden de kans om de niches voor grote landdieren te gaan innemen. Al snel namen verschillende groepen zoogdieren in lichaamsgrootte toe en de placentale zoogdieren begonnen ook de lucht en de zee als leefgebied te gebruiken. Dit leidde tot de huidige situatie waarin er een grote vormenrijkdom aan synapside soorten is met een enorme variatie in lichaamsbouw. Het grootste bekende dier uit de wereldgeschiedenis, de blauwe vinvis, is een synapside en ook de eerste technologische soort, Homo sapiens.
- Enkele synapsiden
-
![Archaeothyris (Eupelycosauria)[2]](//upload.wikimedia.org/wikipedia/commons/thumb/3/36/Archaeothyris_BW.jpg/120px-Archaeothyris_BW.jpg)
-

-

-

-

-

-

-

![Archaeothyris (Eupelycosauria)[2]](?x=http://nl.wikipedia.org/wiki/Bestand:Archaeothyris_BW.jpg)






Fylogenie
[bewerken | brontekst bewerken]Een traditionele indeling van de Synapsida is de volgende:
- Amniota
- Onderklasse Synapsida
- Orde Pelycosauria
- Familie Eothyrididae
- Familie Ophiacodontidae
- Familie Varanopidae
- Familie Sphenacodontidae
- Familie Edaphosauridae
- Familie Caseidae
- Orde Therapsida
- Suborde Biarmosuchia
- Suborde Anomodontia
- Suborde Dinocephalia
- Suborde Gorgonopsia
- Suborde Dicynodontia
- Suborde Therocephalia
- Suborde Cynodontia
- Orde Pelycosauria
- Onderklasse Synapsida
Een mogelijke fylogenetische indeling wordt gegeven in dit kladogram, waarbij de zoogdieren deel zijn van de Synapsida en geen rangen gebruikt worden:
| Synapsida |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cynodontia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ontdekkingsgeschiedenis
[bewerken | brontekst bewerken]
Synapsiden zijn de mens natuurlijk van oudsher bekend daar hij zelf een synapside is en veel van de grotere dieren waarmee hij de planeet deelt, zoogdieren zijn en dus ook weer synapsiden. Dat er een groep bestaat die groter is dan de zoogdieren alleen doordat die uitgestorven directe verwanten bezitten, is een vrij recente ontdekking. De eerste basale Synapsida werden in het begin van de negentiende eeuw in Rusland gevonden, in de Oeral in lagen uit het Perm en in 1838 gemeld door professor Stephan S. Koetorga die dat jaar de geslachten Brithopus en Syodon benoemde. Dergelijke vormen werden echter niet als heel verschillend van moderne zoogdieren beschouwd. Koetorga zag Brithopus als een verwant van de luiaards en Syodon als een oude vertegenwoordiger van de olifanten. In werkelijkheid waren ze zo volstrekt anders gebouwd dat het meestal nog als een hele prestatie van Koetorga wordt gezien dat hij überhaupt een verwantschap met de zoogdieren onderkende.
De Duitse geoloog Friedrich Wangenheim von Qualen onderzocht het gebied kort daarop, naar kopererts speurend, en trof botten van "sauriërs" aan. Dat trok de aandacht van Johann Gotthelf Fischer von Waldheim die in 1841 Rhopalodon benoemde. Tegelijkertijd was de Brit Roderick Murchison bezig de stratificatie van de Oeral te bepalen en groef ook weer fossielen op die hij doorstuurde naar Richard Owen in Engeland. Owen zag er, zoals zo vaak, meer een krokodilachtige in. Dat veranderde toen er een tweede belangrijk vondstgebied van basale synapsiden ontdekt werd: Zuid-Afrika waar in 1845 Andrew Geddes Bain fossielen van therapsiden meldde. In 1876 begreep Owen dat het om vroege voorlopers van de zoogdieren moest gaan. In 1877 werd een volgende belangrijke stap gezet toen Edward Drinker Cope als eerste de link legde tussen de Zuid-Afrikaanse therapsiden en Amerikaanse vondsten van "Pelycosauria", waarvan de eerste, Bathygnathus, in 1854 door Joseph Leidy benoemd was zonder dat die de speciale aard daarvan onderkende. Die gedachte werd echter door lang niet alle onderzoekers aanvaard. Daar kwam in het begin van de twintigste eeuw verandering in door het werk van de Schot Robert Broom die in Zuid-Afrika talloze fossielen opgroef. Hij benoemde de Therapsida formeel en maakte een streng onderscheid met de eerdere en "primitievere" Pelycosauria, een schema dat tot ver in de twintigste eeuw gangbaar zou blijven.
Tegen het eind van de twintigste eeuw maakte de methode van de cladistiek het mogelijk de verwantschappen tussen diergroepen exact te berekenen. Daaruit bleek dat de "Pelycosauria" een parafyletische groep waren, een reeks opeenvolgende afsplitsingen van de hoofdstam van de Synapsida. Die werden nu apart als kladen gedefinieerd en het verouderde begrip Pelycosauria werd verlaten. Dit proces viel samen met een constante stroom aan nieuwe vondsten die leidden tot een sterk vergroot inzicht in de bouw van de synapsiden.
Beschrijving
[bewerken | brontekst bewerken]Onderscheidende kenmerken
[bewerken | brontekst bewerken]Zoals alle diergroepen hebben synapsiden synapomorfieën, nieuwe kenmerken die ze delen. Het belang dat Osborn hechtte aan de slaapvensters, wordt door modern onderzoek gerechtvaardigd. Het bezit van alleen een onderste slaapvenster aan de zijkant van de schedel blijkt nog steeds een geldige synapomorfie te zijn. Daarbij moet bedacht worden dat er bij latere zoogdieren in hun volwassen vorm van alles met dat slaapvenster kan gebeuren, tot volledige sluiting aan toe. Dat maakt het niet minder tot een synapomorfie, waarvoor het criterium is of men binnen de groep ooit een voorouder heeft gehad met het kenmerk. Het is de vraag wat de functie van het slaapvenster is; meestal wordt aangenomen dat het voordeel ligt in een combinatie van vergroting van het aanhechtingsvlak van de sluitspieren, gewichtsbesparing en energiebesparing door het niet hoeven op te bouwen en onderhouden van beenweefsel buiten de essentiële krachtlijnen. Overigens is het in het kader van de tegenstelling met de Diapsida relevanter waarom synapsiden geen bovenste slaapvenster ontwikkelden, iets waarover weinig inzicht bestaat.
Andere mogelijke synapomorfieën zijn het bezit van een enkelvoudig os postparietale op de middenlijn van het schedeldak; de begrenzing bovenaan van het slaapvenster door het postorbitale en het squamosum; een breed en naar voren hellend achterhoofd; en een septomaxilla — het bot tussen de maxilla ofwel bovenkaaksbeen en de praemaxilla — met een brede basis en een massief gebouwde opgaande tak.
Skelet
[bewerken | brontekst bewerken]Schedel en onderkaken
[bewerken | brontekst bewerken]
De evolutie van schedel en onderkaken verliep bij de Synapsida langs heel andere lijnen dan bij de Diapsida. In die laatste groep was de algemene trend er een van steeds lichtere schedels met meer en grotere vensters. Vaak was de verbening onvolledig en ook ontwikkelden zich in veel gevallen gewrichten tussen de schedelelementen zodat er een kinesis, onderlinge bewegelijkheid, mogelijk werd. Snuit en kaken konden ten opzichte van de hersenpan allerlei strekkingen en buigingen uitvoeren.

Bij de Synapsida daarentegen was er een trend naar steeds zwaardere schedels. Het onderste slaapvenster waarna de groep genoemd is, werd zo allerminst een aanzet tot grotere beweeglijkheid. Het werd gebruikt voor de aanhechting van steeds zwaardere sluitspieren. De belangrijkste spiergroepen liepen niet meer van de onderkant van het verhemelte naar de binnenkant van de onderkaak maar over de buitenkant van de slaap naar de buitenkant van de onderkaak. Om dit alles te verstevigen vergroeide het vierkantsbeen met de hersenpan en werd het achterhoofd een brede plaat. In een latere fase werd het schedeldak smaller door reductie van het posttemporale en het postorbitale zodat een belangrijker deel van de spierkracht verticaal gericht kon worden wat de beet versterkte. Tegelijkertijd kreeg de onderkaak achteraan een hoger coronoïde uitsteeksel voor een betere hefboomwerking, terwijl het articulare, dat het onderste kaakgewricht vormde, juist lager kwam te liggen om dit effect te versterken. Een naar onderen vergrote beenplaat aan de onderste achterkant van de onderkaak, het angulare, verhoogde het aanhechtingsvlak. Door dit alles werd de onderkaak achteraan heel hoog. Daarbij ontstond er een ruimte tussen de kiel van het angulare en het articulare, in de onderzijde van de kaak. Daarin hechtte zich een extra spiergroep.
Vroege therapsiden erfden dit systeem. Het had het nadeel dat bij de vrij lange koppen die synapsiden hadden de statische bijtkracht bij de lange slagtanden vooraan tamelijk gering moet zijn geweest. Het is wel aangenomen dat dit gecompenseerd werd door een vermogen de muil zeer sterk te openen zodat de slagtanden bij sluiting lang konden versnellen om met een grote vaart dicht te klappen zodat de huid van een prooi toch doorboord kon worden. Achteraan werd bij de Therapsida de spierkracht vergroot door de ontwikkeling van een teruggebogen beenplaat van het angulare wat extra ruimte schiep voor de musculatuur. Het is echter ook wel geopperd dat die diende voor een klier of in verband stond met de latere rol van het angulare in het gehoor. Mochten hier inderdaad spieren aangehecht zijn, is het niet zeker of het om sluitspieren gaat of juist om een spier die naar de hals liep, om de muil te openen. Ook tongspieren kunnen hier geplaatst zijn.
Bij latere therapsiden deed zich een enorme vergroting van het slaapvenster voor, naar boven en binnen toe. Het lag daardoor meer horizontaal en besloeg niet alleen bijna de hele achterste zijkant van de schedel maar ook het grootste deel van het schedeldak. Tussen beide slaapvensters lag middenin nog een smalle kam. Dit diende voor een aanzienlijke verbreding van het interne deel van de sluitspier, de Musculus adductor mandibulae internus. Hierdoor kwam een spier die eigenlijk aan de binnenkant van de schedel lag bovenaan meer aan de buitenzijde te liggen; bij zoogdieren wordt hij daarom de Musculus temporalis genoemd. Het jukbeen, de oude onderste zijkant van de schedel, lijkt hierdoor een aparte boog, een "juk". Voor een betere hefboomwerking van de spier schoof het coronoïde uitsteeksel van de onderkaak naar voren, het os dentale op. Door deze verschuiving werd de achterste tak van de onderkaak bij sluiting meer naar achteren en binnen getrokken. Dit werd opgevangen door aanpassingen aan het vierkantsbeen, het quadratum. Het kwam onderaan meer schuin naar voren te staan en werd ook bovenaan losser verbonden aan het squamosum zodat het om zijn lengteas kon draaien. Zo kon het meebewegen met de achterste tak van de onderkaak zodat een articulatie van het kaakgewricht mogelijk bleef. De kaak kon de trekkracht niet meer naar voren afleiden want de onderkaken waren vooraan hecht verbonden zodat er een enkele mandibula gevormd werd.
Basale cynodonten hadden met hun ingewikkelder gebit en beginnend vermogen tot kauwen meer spierkracht nodig. Daartoe verlegde de Musculus temporalis zich verder naar beneden op de onderkaak, naar de groeve van de fossa adductoris. Verder ontwikkelde zich een spier van de jukboog naar de onderkaak, de Musculus masseter. Deze trok de kaak naar boven en buiten. Nu er twee zware spieren waren die de kaak naar binnen en naar buiten trokken, kon de kauwbeweging overdwars veel beter beheerst worden. Daarentegen verminderde het teruggebogen blad van het angulare in omvang. Door de werking van de masseter, werd het vermogen tot rotatie van het vierkantsbeen ook minder belangrijk.
Bij de Epicynodontia deed zich een volgende ontwikkeling voor: het coronoïde uitsteeksel van het os dentale raakte verticaal verlengd en begon nu diep in het slaapvenster te steken, van onderen uit, achter de jukbeenboog langs. Het werd bijna net zo hoog als de schedel zelf. De jukbeenboog werd veel groter en raakte bovenaan gewelfd voor een vergrote masseter die nu naar beneden doorliep over een nieuw ontwikkelde onderste beenplaat, het angulare in functie vervangend, van het os dentale. De verticale krachten op de onderkaak kwamen zo meer naar voren te liggen, weg van het kaakgewricht. Tegelijkertijd ontwikkelde het surangulare van de onderkaak een knobbel voor een kapsel dat het met het squamosum verbond, het begin van een nieuw kaakgewricht. De nieuwe situatie met grote opgaande tak schept de illusie dat zich daar dat nieuwe gewricht begon te ontwikkelen maar het zou meer achteraan gaan liggen. Door vroege paleontologen werd de opgaande tak bij vroege vormen wel aangezien voor een deel van het cranium.
Bij de Eucynodontia bereiken al deze processen hun logische eindpunt. De opgaande tak van het uitsteeksel van het os dentale wordt even hoog als de schedel zelf, het os dentale begint naar achteren groeiend zelf het kaakgewricht te vormen, de achterste onderkaaksbeenderen raken sterk gereduceerd en de krachten op de onderkaak worden zo gebalanceerd dat een beweeglijk vierkantsbeen daarvoor niet meer nodig is en de grootste bijtkracht ontstaat waar de bovenste en onderste tanden elkaar in occlusie raken. De reductie van de achterste beenderen bevrijdde deze voor een verbetering van de gehoorfunctie. De Tritylodontidae laten zien dat het verlies van de postorbitale beenbalk vooraf ging aan de vorming van een nieuw kaakgewricht tussen het os dentale en het squamosum. Bij de Triheledonta is dit kaakgewricht nog erg simpel, zonder knobbel op het os dentale of groeve in het squamosum. De onderste tanden beginnen nu de binnenzijde van de bovenste tandrij te raken maar daarvoor werden de onderkaken heen en weer bewogen. De Mammaliamorpha hebben wel een knobbel op het dentarium die in een squamosale holte past. Het oude achterste gewricht is er eerst nog maar de functie ervan is overgenomen. Het meer robuuste voorste gewricht kan de horizontale krachten opvangen van de eerst nog simpele horizontale occlusie.
Tanden
[bewerken | brontekst bewerken]De tanden van de Synapsida leken in eerste instantie veel op die van reptielen. Iets waarin ze zich al vroeg onderscheiden, is een trend naar heterodontie: het voorkomen van verschillende typen tanden in de kaken, vooral grotere hoektanden naast ronde grijptanden vóór in de kaken. De Anomodontia, die zo heten vanwege hun afwijkende gebit, verloren die hoektanden maar ontwikkelden een nieuw soort slagtanden daarvoor in de plaats. Waarin ze langer met de reptielen overeenkwamen, is de polyphyodontie: het continu vervangen van de tanden door de oude af te stoten door het groeien van onderliggende vervangingstanden. Er was een doorlopende cyclus waarin tanden van achteren naar voren in de rij vervangen werden. Dat leidde onherroepelijk tot een onregelmatig gebit en daardoor een slechte occlusie. Dat belemmerde weer een efficiënte kauwwerking en paste ook slecht bij de heterodontie met haar duidelijke afgescheiden sectoren in de tandrij. Bij basale Cynodontia ontwikkelde zich daarom een systeem van diphyodontie: de tanden worden maar eenmaal gewisseld en na een melkgebit verschijnt het volwassen gebit. Het nieuwe systeem had ook nadelen. Het verlies van een volwassen tand zal het gebit onherstelbaar beschadigen. Bij wat kleinere cynodonten maakte dat echter weinig uit omdat die toch maar een korte levensverwachting hadden. Voor de groter evoluerende zoogdieren werd dit echter tijdens het Kenozoïcum snel een probleem. Vaak sterven die op hogere leeftijd omdat hun gebit afgesleten is. Slechts kleine groepen konden dit ontlopen door helemaal tandeloos te worden. De anomodonte dicynodonta lieten hun individuele slagtanden continu aangroeien. Knaagdieren deden later hetzelfde met hun snijtanden, een verschijnsel dat hypselodontie heet.
Bij latere groepen Synapsida ontwikkelde de heterodontie zich verder in een typische verdeling in snijtanden, hoektanden, premolaren en kiezen waarbij iedere tandsector zijn eigen functie had. De precieze indeling en vorm van de tanden verschilde hierbij sterk van groep tot groep, zodanig dat men uitgestorven soorten al aardig kan determineren op grond van één gevonden tand. Bij basale cynodonten deed zich voor het eerst een onderscheid tussen premolaren en kiezen voor, waarvan ze er per kwadrant een dozijn hadden. Het onderscheid was echter nog niet heel sterk zodat men wel spreekt van "premolariforme" en "molariforme" tanden. Die laatste kregen geleidelijk een kiesvorm. Eerst ontwikkelden ze een cingulum, een opstaande rand rond de basis, en daarop kwamen uiteindelijk kleine knobbeltjes die bijvoorbeeld handig waren om harde insectenpantsers te kauwen. In deze fase was er nog geen precieze occlusie maar wel raakten tanden van bovenkaken en onderkaken elkaar. Bij eucynodonten kregen de tanden een "triconodonte" vorm met een middelste hoofdpunt geflankeerd door twee lagere punten.
Wervelkolom
[bewerken | brontekst bewerken]Niet-therapside synapsiden hadden typisch zo'n tachtig tot negentig wervels. Ze hadden vijf tot zeven halswervels en achttien tot twintig ruggenwervels voor een totaal van vierentwintig tot zevenentwintig presacrale wervels. Het aantal sacrale wervels in het heiligbeen bedroeg twee, drie of vier. De staart telde vijftig tot vijfenzestig wervels, soms boven de zeventig. Basale therapsiden toonden tot drieëndertig presacralen waaronder tot acht halswervels. Hun staart was vaak verkort, soms tot vijftien wervels. Basale cynodonten konden tot achtentwintig presacralen hebben, negen halswervels, vier of vijf sacrale wervels en weer een verkorte staart, soms met niet meer dan tien staartwervels.
De basale Synapsida hadden alle dezelfde vorm van de eerste twee halswervels. In het contact met de schedel werd voorzien door bovenaan een gepaarde proatlas en onderaan een intercentrum van de atlas. De atlas zelf bestond uit een vergroeid pleurocentrum met daarop een onvergroeide gepaarde wervelboog. Daarachter kwam onderaan een intercentrum van de draaier en de draaier bestond weer uit een vergroeid pleurocentrum met een vergroeide wervelboog. Bij de Therapsida werd de enkelvoudige achterhoofdsknobbel verbreed tot een niervorm. Daardoor werd een schuddende beweging van de kop mogelijk. Iedere deelknobbel raakte een stuk van de gepaarde wervelbogen van de atlas, wat ook het knikken vereenvoudigde.
Het atlas-draaier-complex werd oorspronkelijk aangevuld met nog eens vier halswervels. Basale Synapsida konden er ook drie hebben, vijf, zes of zeven, vaak nog met intercentra ertussen. Die werden bij basale Therapsida zelfs dikker wat de beweeglijkheid van de nek vergrootte. De huidige zoogdieren hebben meestal zeven halswervels in totaal. De variabiliteit daarin is dus gering wat een scherp contrast vormt met de Diapsida waarvan sommige vormen wel tweeënzeventig halswervels bezaten. Dergelijke fundamentele verschillen zijn een uitdrukking van onderliggende hoxgenen. Basale Synapsida hadden nog zijuitsteeksels in de halswervels en nekribben. Huidige zoogdieren hebben hun nekribben verloren.
Al bij niet-therapside synapsiden moet de horizontale beweeglijkheid van de ruggenwervels verminderd zijn zoals blijkt uit het feit dat hun gewrichtsuitsteeksels niet meer horizontaal gewricht zijn maar schuin. Eveneens worden de intercentra klein. Vormen met een rugzeil bogen dat vermoedelijk niet meer horizontaal. Hun ribben bleven echter beweeglijk en vormden de voornaamste ondersteuning van de romp. Bij de Therapsida vallen de intercentra in de rug helemaal weg. Bij basale vormen zijn de gewrichtsuitsteeksels ook verticaal georiënteerd wat een horizontale beweeglijkheid vrijwel uitsluit. Ook verticaal was de ruggengraat echter nog vrij stijf.
Bij de Therocephalia beginnen de achterste ruggenwervels hun ribben te verliezen zodat er een onderscheid kan worden gemaakt tussen borstwervels en lendenwervels.
Niet-therapside synapsiden hebben meestal nog buikribben of gastralia, een korf vormend op de onderzijde van de romp. Bij de Therapsida heeft nog maar een enkele soort buikribben die bij cynodonten niet meer worden aangetroffen. Hun verdwijnen kan verband houden met de vorming van een middenrif.
Ledematen
[bewerken | brontekst bewerken]De schoudergordel had bij de eerste Synapsida een interclaviculum met een ruitvormige kop. Het procoracoïde was oorspronkelijk groter dan het ravenbeksbeen; bij meer afgeleide therapsiden werden ze even groot. Bij niet-therapside synapsiden vormde de schoudergordel een massief stijf juk op de voorste borstkas met een groot schouderblad bovenop en een coracoïde bodemplaat onderop. Het schoudergewricht had een schroefvorm: van voor naar achter roteerde het gewrichtsvlak van beneden/achterwaarts via zijwaarts naar boven/voorwaarts. Bij therapsiden raakten de sleutelbeenderen los van de lichtere schoudergordel die nu ten opzichte van de ruggengraat kon bewegen om de paslengte van de voorpoot te verlengen.
Het opperarmbeen had bij niet-therapside synapsiden bovenaan een spiraalvorm om in het schoudergewricht te passen. Bij het begin van de pas stak het naar achteren en aan het eind naar voren. Bij basale therapsiden werd de schouder een zadelgewricht en de kop van het opperarmbeen werd cilindervormig zodat het in dat gewricht een rolbeweging kon uitvoeren. Hierdoor was het mogelijk de romp tijdens de pas meer te heffen. Het opperarmbeen had bij de meest basale synapsiden een sterk verbreed onderste uiteinde met krachtige beenstijlen. Het had oorspronkelijk een sterk gewrongen schacht. De ellepijp had eerst een sterk verlengde processus olecrani, wat de spreidstand van de voorpoten weerspiegelt. De pols had oorspronkelijk twee carpalia voor ieder middenhandsbeen.
Bij niet-therapside synapsiden had het bekken een simpele plaatvorm. Rotatie van het onderbeen vond plaats bij de knie. Bij basale therapsiden werd de heup een balgewricht. Zo kon de achterpoot zowel zijdelings als verticaal gehouden worden. Hiertoe moest de enkel flexibeler worden en daartoe evolueerde een intern enkelgewricht tussen sprongbeen en hielbeen, een ontwikkeling die haar parallel heeft bij de latere Crurotarsi. Niet langer de staartspieren maar de bilspieren en de Musculus iliacus waren de belangrijkste retractors.
De synapsiden hadden oorspronkelijk naar alle waarschijnlijkheid dezelfde formules voor de kootjes als bij de eerste Amniota, namelijk 2-3-4-5-3 in de hand en 2-3-4-5-4 in de voet. Bij de zoogdieren werd dat 2-3-3-3-3 voor zowel hand als voet.
Levenswijze
[bewerken | brontekst bewerken]Habitat
[bewerken | brontekst bewerken]Het typische leefgebied van de eerste synapsiden werd tijdens het Carboon gevormd door de tropen van het zich ontwikkelende supercontinent Pangea. Het klimaat was heet en vochtig en de habitat kende als tropische zone geen seizoenen. De flora bestond grotendeels uit paardestaarten, wolfsklauwen en varens, hoewel coniferen in opkomst waren. De planeet als geheel was echter nog niet zo heet: er waren ook gematigde zones en op het Zuidelijk Halfrond lag een grote ijskap. Buiten de toenmalige tropen zijn echter geen basale synapsiden aangetroffen. Het weelderige plantendek had veel voedsel voor grote planteneters kunnen opleveren maar die waren er maar weinig. Caseïden en edaphosauriden waren de uitzondering en zullen maar een klein beslag gelegd hebben op de aanwezige biomassa. De planten werden gegeten door ongewervelden en die weer door insecteneters, waaronder maar weinig synapsiden en veel "amfibieën" — dus niet-amniote tetrapoden. Een andere goede niche was die van viseter en daarin waren de ophiacodontiden gespecialiseerd naast opnieuw veel "amfibieën". Aan de top van de voedselketen stonden die dieren die weer op de insecteneters en viseters joegen, een rol die de sphenacodontiden vervulden.
In de loop van het vroege Perm werden de omstandigheden steeds droger en de meeste habitats kenden nu een droog seizoen. De edaphosauriden werden talrijker en daarmee ook de sphenacodontiden die van ze leefden. Volgens Olson maakten de caseïden een nog grotere groei door en werden die bejaagd door varanopiden maar Thomas Kemp wees erop dat het bewijs hiervoor gebaseerd was op één locatie in Oklahoma. In ieder geval vormden de synapsiden de eerste fauna van grote gewervelde landdieren die niet direct afhankelijk was van de productiviteit van zoetwateroppervlakken. "Amfibieën" werden veel minder belangrijk. De ontwikkeling van rugzeilen wijst mogelijkerwijs op een wat actievere levenswijze.

De overgang van het vroege Perm naar het middelste Perm werd niet gekenmerkt door abrupte klimaatsveranderingen of verschuivingen van de continentale platen. Wel smolten de ijskappen af en werden zo veel bredere gematigde zones productief, begroeid door zaadvarens. Dit werd kennelijk volop geëxploiteerd door de therapsiden die na Olson's Gap opduiken. Die waren nog actiever. De schedels van vleeseters werden zwaarder, met grotere slagtanden en krachtiger sluitspieren. Planteneters ontwikkelden complexere gebitten, om meer soorten voedselplant meer efficiënt te kunnen verwerken. Beide categorieën evolueerden langere, slankere, soepelder en rechtere ledematen, voor vlucht en achtervolging over grotere afstand. Hun stofwisseling ging omhoog. Daardoor konden ze ook koudere gebieden bevolken en hun hogere energiebehoefte deed ze een groter beslag leggen op het beschikbare voedsel zodat ze de meer basale synapsiden wegconcurreerden. Zelfs de caseïden, die zich specialiseerden in een geringe voedselinneming, stierven uit. Dit is een van de zeldzame gevallen waarvoor het ook naar moderne inzichten plausibel geacht kan worden dat op wereldschaal een nieuwe fauna de oude verving vanwege een algemene superioriteit in "organisatie". De therapsiden beconcurreerden ook zichzelf en zo was er een snelle afwisseling in soorten, waarbij hele deelgroepen her loodje legden en vervangen werden. Er bleef echter ook een rol weggelegd voor wat grotere plantenetende koudbloedige reptielen en de therapsiden deelden hun leefgebied met de Pareiasauria die zich verdedigden door pantsering. In het water leefden amfibische Temnospondyli.
De catastrofe op het eind van het Perm verwoestte het plantendek met als gevolg een enorme erosie en woestijnvorming. Slechts een dozijn synapside afstammingslijnen schijnt het te hebben overleefd. Lystrosaurus was direct daarna wellicht daarom succesvol omdat hij goed kon graven en zo knollen en wortels bereiken. Daarna herstelden de planten zich weer en er ontstond nu een warm klimaat met een duidelijke seizoenswisseling. Hoewel dat niet fundamenteel verschilde van de situatie in het late Perm, wisten de therapsiden zich niet helemaal te herstellen. Ze kregen steeds meer concurrentie van Archosauromorpha zoals de plantenetende Rhynchosauria. Na het eind van het Carnien, zo'n 220 miljoen jaar geleden, ging het hard met de neergang van de synapsiden. Eerst verdwenen de grotere rovers en uiteindelijk ook de grote planteneters. Vaak is dit geweten aan concurrentie met de opkomende dinosauriërs. Er kunnen ook een of meer massa-extincties geweest zijn maar die roeiden niet alle synapsiden uit zodat alleen een "superioriteit van de dinosauriërs", zoals Robert Thomas Bakker het uitdrukte, schijnt te kunnen verklaren waarom die dominant bleven. Op het einde van het Trias waren alle synapsiden kleine diertjes met een beperkte rol in hun ecosysteem.
Voortbeweging
[bewerken | brontekst bewerken]De eerste synapsiden bewogen zich voort als basale Amniota: door een zijdelings golvende beweging van het lichaam. De poten stonden daartoe zijwaarts. De Therapsida gingen hoger op de poten staan die onder het lichaam stonden. De horizontale golvende beweging ging verloren en opperarmbeen en dijbeen draaiden niet meer om hun lange as. De beweging van de ledematen van voor naar achter werd versterkt alsmede de strekking van handen en voeten. Tegelijkertijd werd de schoudergordel losser en kon de ruggengraat verticaal buigen zodat de voorpoten beter de achterpoten konden bijhouden die echter wel het grootste deel van de voortstuwende kracht bleven leveren. De basale therapsiden konden zich zowel laag en langzaam door de poten gezakt voortbewegen als overschakelen naar een snellere voortbeweging hoog op de poten.
Zie ook
[bewerken | brontekst bewerken]Noten
- ↑ Lambert, David (2001). Dinosaur Encyclopedia. ISBN 0-7894-7935-4. pp. 68–69.
- ↑ De groep waar een dier in deze plaatjesgalerij bij ingedeeld is, omvat ook alle dieren die op de volgende plaatjes te zien zijn
Literatuur
- H.F. Osborn. 1903. "On the primary division of the Reptilia into two sub-classes, Synapsida and Diapsida". Science 17(424): 275-276
- Kemp, T.S. 1982. Mammal-Like Reptiles and the Origin of Mammals. Academic Press, London
- Hotton, N., MacLean, P.D., Roth, J.J. & Roth, E.C. 1986. The Ecology and Biology of Mammal-like Reptiles. Smithsonian Institution Press, Washington and London
- Robert R. Reisz, 2004, "Early Amniote Phylogeny and Nomenclature", First International Phylogenetic Nomenclature Meeting, Paris, Muséum National d’Histoire Naturelle, July 6-9, 2004 Under the auspices of the Académie des Sciences de Paris, Abstracts p. 25
- Angielczyk, K.D. 2009. "Dimetrodon is not a dinosaur: using tree thinking to understand the ancient relatives of mammals and their evolution". Evolution: Education and Outreach 2: 257-271
- Roger B. J. Benson, 2012, "Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies", Journal of Systematic Palaeontology, 10(4): 1-24 DOI:10.1080/14772019.2011.631042